3.1. Общие закономерности усиления превентивно-защитной вегетативной активности при стрессе
Если взглянуть на вегетативные процессы, не подчиненные прямым влиянием воли человека, оценивая их сопряженность с психическими процессами, то можно увидеть три типа взаимоотношений вегетатики с психикой (см. рис. 15). 1) Вегетатика "обслуживает" психическую сферу. Психические процессы базируются на физиологических механизмах. При этом психика как бы "управляет" вегетатикой. Их взаимодействие призвано обеспечивать оптимальный режим психической деятельности. 2) В некоторых случаях, в частности при стрессе, вегетатика может принимать на себя функции, которые с определенным допущением можно расценивать не только как "обслуживание" психических процессов, но и как "управление" ими. Например, в сильную жару происходит выбрасывание из организма с мочой многих активаторов мышечной и психической активности; у людей возникают чувства мышечной слабости, апатия, нежелание двигаться. Известно, что уменьшение объема движений способствует защите организма от перегрева. В данном случае "обеспечение" оптимального, т. е. минимального, объема и интенсивности движений происходит за счет снижения мышечной силы и желания двигаться, за счет возникновения апатии. Можно привести много примеров участия вегетативных механизмов в регуляции поведения. Следует заметить, что "высшая власть" сознания и воли человека в регуляции своего поведения даже в критических ситуациях, при стрессе, сохраняется. Вопрос в том, в какой степени человек сможет воспользоваться этой "властью", т. е. насколько он знает, что ему надо делать в тех или иных экстремальных условиях, насколько он умеет это делать и насколько социальное окружение поможет ему в трудную минуту подчинить свое поведение воле и разуму. 3) Вегетатика может выполнять "защитные" функции. При стрессе они могут резко усиливаться, становясь весьма заметными для субъекта и для окружающих. В ожидании опасности, т. е. превентивно, могут усиливаться те или иные, иногда многие формы защитной вегетативной активности [129, 130, 158 и др.].
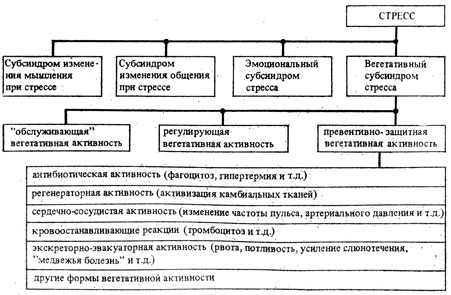
Рис. 15. Схема развития вегетативного субсиндрома стресса
Рассмотрим подробнее взаимосвязь поведения и защитной вегетативной активности при стрессе, а также индивидуальные особенности защитных вегетативных реакций. В первых же исследованиях острого стресса в невесомости было обнаружено, что у испытуемых, которые реагировали на экстремальный фактор снижением активности поведения (пассивное эмоционально-двигательное реагирование), одновременно возникали своеобразные пространственные иллюзии. Таким людям казалось, что сила тяжести не исчезает, как это происходило в действительности, а начинает действовать в противоположном направлении, т. е. возникает "тяга вверх", человека будто бы тянут туда, куда обращено его темя [113, 114]. Эта иллюзия была интерпретирована как результат ряда процессов, в частности актуализации в сознании противообраза исчезнувшего действия тяжести [123, 126]. Было непонятно, почему повторение режимов невесомости вызывало негативные "расстройства" по типу морской болезни только у людей этой группы. У лиц с активными эмоционально-двигательными реакциями при кратковременной невесомости таких "расстройств" не наблюдалось. Было предложено следующее объяснение этого феномена. Ситуация, порождающая иллюзорное представление о "тяге вверх", тогда как тела человека не касаются никакие предметы, представляется ему невозможной (на неосознаваемом уровне). Возникает большая субъективная вероятность (субъективная определенность) сигнала, невозможного с позиции фило- и онтогенетического опыта. В фило- и онтогенетической памяти субъекта, естественно, нет программы защитного поведения, адекватного "невозможной" ситуации. Наличие значимого сигнала требует реагирования на него, отсутствие специфических программ защитного поведения препятствует выбору из фило- и онтогенетически опробованных реакций подходящей случаю формы поведенческого реагирования. Тем не менее защитное реагирование осуществляется. Это, прежде всего, отказ от какого-либо активного поведения, т. е. пассивное эмоционально-двигательное реагирование. Оно, как указывалось, характеризует людей, отнесенных ко второй группе. Пассивное реагирование - это не пассивный отдых. Оно приводит к расходу адаптационных резервов. Угроза их исчерпания требует если не поведенческих, то каких-то иных форм активной защиты. Поэтому спустя некоторое время вместо "выключенной" поведенческой активности включается вегетативная защитная активность. Она возникает и тогда, когда имеется лишь опасность еще не начавшего действовать вредоносного фактора. В таком случае она превентивно-защитная. Можно привести ряд примеров того, как явления, невероятные с позиции личного опыта (вид необычного уродства, расчлененного человеческого тела, неожиданный сильный грохот, надвигающаяся неясная опасность и т. п.), могут вызвать рвоту, холодный пот, непроизвольное мочеиспускание и другие виды стрессогенной превентивно-защитной вегетативной активности.
Выше указывалось, что кратковременный стрессор вызывает пассивное поведенческое реагирование (первичное) и сопровождающую его вегетативную защитную активность только у части популяции (у другой ее части возникает стрессовая активизация поведения). В отличие от этого длительный непрерывный или прерывистый стрессор практически у всех людей, на которых он действует, вызывает пассивное поведенческое реагирование (вторичное), также сопровождающееся симптомами активизации вегетативной "защиты". Как первичное, так и вторичное пассивное поведение формируется при "субъективной невозможности" текущей ситуации. Если в первом случае информация о такой "невозможности" порождается отсутствием в фило- и онтогенетическом опыте субъекта прецедентов столкновения со стрессорами, аналогичными тому, который действует в текущий момент, то во втором случае происхождение сигналов о невозможности ситуации иное.
Для анализа возникновения этой информации рассмотрим изменения при повторяющихся стрессорах (при экстремальной монотонии) "комплекса" активного реагирования на одиночный стрессор (см. выше). Этот комплекс, т. е. защитная реакция (действие), призван, условно говоря, не допускать повторного возникновения экстремального воздействия. При монотонной стимуляции каждый новый экстремальный стимул вновь вызывает "включение" такого комплекса. В результате оказывается, что при непрекращающейся экстремальной стимуляции эти защитные комплексы активного реагирования не предотвращают повторного поступления стрессогенных стимулов. Накапливается информация (как правило, неосознаваемая) о неэффективности использования в качестве защиты той формы активного эмоционально-двигательного реагирования, которая реализуется при каждом комплексе. Ситуация с одиночным стимулом, воспринимаемая (на неосознаваемом уровне) как определенная и возможная, условно говоря, "понятная" субъекту, оказывается при монотонной стимуляции невозможной с позиции фило- и онтогенетического опыта, как бы "непонятной". Результат этого - "дискредитация" программ активного реагирования и замена его пассивным реагированием. Этот процесс усугубляется, если последующий стимул действует до завершения "комплекса" активного реагирования на предыдущий стимул. Это препятствует завершению "комплекса", деформируя и разрушая его экстатическую фазу, что, в свою очередь, нарушает восстановление физиологического и психологического гомеостаза.
Экстремальный стимул - это первичный стрессогенный фактор. Накопление информации о неуспешности "комплексов" активного реагирования и о незавершенности гомеостатических процессов становится вторичным экстремальным, стрессогенным фактором. Усиливается мобилизация глубоких адаптационных резервов и перестройка "функциональной системности" организма. При этом на смену более специализированной двигательной, поведенческой адаптационной активности приходит усиление вегетативной активности с широким фронтом "защиты". Имевшееся в начале развития стресса доминирование эмоционального субсиндрома стресса сменяется доминированием вегетативного субсиндрома.
При длительном действии прерывистого стрессора были прослежены различные формы вегетативной превентивно-защитной активности. Каковы эти формы? Почему их можно назвать защитными? В отличие от защитного поведения (агрессии или бегства), которое у человека в той или иной мере нацелено сознанием на удаление конкретного вредоносного фактора, вегетативная защита, непосредственно не контролируемая сознанием, не может иметь такой "нацеленности". Это в некоторой степени компенсируется "многоканальностью" вегетативной защиты, набором защитных реакций, "на всякий случай". Рассмотрим следующую гипотетическую структуру возникновения основных форм таких реакций.
Усиление экскреторно-эвакуаторной защитной функции. Допустим, в интегративных центрах нервной системы, на неосознаваемом уровне актуализуется концепт того, что неясная опасность - это яды, проникшие в организм, или токсические метаболиты. Решение - их надо выбросить. Результат - усиление экскреции жидкости из организма и эвакуация из его полостей. Возрастают пото-, слюно-, мочевыделение. Увеличивается секреция желез слизистой оболочки желудка и кишечника. Содержимое последних выбрасывается благодаря антиперистальтике желудка и двенадцатиперстной кишки и усилению перистальтики толстого кишечника и прямой кишки. Возникают тошнота, рвота и "медвежья болезнь".
Усиление гемоциркулярной функции. Концептуальная модель: "неясная", условно говоря, опасность может стать "ясной" и потребовать перехода от пассивного пережидания к активному бегству или нападению. Решение: превентивно усилить кровоснабжение органов и тканей организма. Результат: учащение и усиление сердцебиения, повышение артериального давления и т. п. Усиление антигеморагической функции. Неясная пока опасность может привести к ранению и кровотечению. Повышается свертываемость крови, возрастает число тромбоцитов и т. д.
Усиление регенераторной функции. Не исключена опасность нарушения целостности тела субъекта и кровопотери. Возрастает активность камбиальных (ростковых) тканей, увеличивается число эритроцитов и других форменных элементов крови. Усиливается антимикробная потенция. На случай опасного инфицирования организма увеличивается продукция форменных элементов белой крови, может резко возрастать температура тела и т. д.
Как указывалось, при остром стрессе только у части из числа обследованных обнаружены превентивно-защитные вегетативные реакции, тогда как при длительном (прерывистом) стрессе такие реакции возникали практически у всех испытуемых. Важным явилось то, что при длительном стрессе эти реакции были не одинаковы у тех, кто при остром стрессе отличался пассивным эмоционально-двигательном реагированием, и у тех, у кого было активное поведенческое реагирование. При длительном гравитоинерционном дистрессе для первых было характерно общее повышение артериального давления, рвота и резкое увеличение пото-, слюно-, а иногда и мочевыделения, т. е. тотальное усиление экскреторно-эвакуаторной функции и т. д. У вторых имели место повышение давления в центральной артерии сетчатки глаза (т. е. реакции церебральных сосудов) и стенокардитические явления (реакция сердечных сосудов) при неизменности общего артериального давления, а также симптомы гиперацидной секреции желудка при отсутствии общей экскреторно-эвакуаторной реакции и т. д. Это можно расценивать как локальные проявления вегетативных реакций, защитных только в своей тотальной форме и теряющих свой защитный модус в локальной форме.
В экспериментах с многосуточным вращением было обнаружено, что дополнительные кратковременные стрессоры при наличии у испытуемых отрицательного эмоционального отношения к ним усугубляли симптомы длительного дистресса (рис. 7, 16, 20, 21). Дополнительные кратковременные стрессоры, вызывающие положительные эмоции, как правило, поначалу уменьшали выраженность имевшихся симптомов дистресса, однако их последействие было разным [129, 130, 158 и др.].
Если такой стрессор вызывал острое чувство ответственности, например при посещении кабины вращающегося стенда высокопоставленным руководителем, выраженность проявлений дистресса первоначально уменьшалась. Однако сразу после такого посещения, то есть как только чувство ответственности за свой внешний вид, за свою работоспособность уменьшалось, происходило резкое возрастание выраженности симптомов дистресса, имевшихся до такого кратковременного эмоционального стрессора. Известны случаи, когда аналогичный кратковременный стрессор, надолго создавая у людей, находившихся в состоянии дистресса, чувство ответственности, оказывал длительное положительное последействие и использовался как один из способов улучшения самочувствия и работоспособности в экстремальных условиях [248, 249, 256 и др.].
Кратковременное эмоциональное, эротически окрашенное переживание также уменьшало выраженность симптомов длительного стресса. Такое переживание могло возникать у испытуемых во время посещения стенда кем-либо из числа врачей, психологов- женщин, если эта женщина вызывала симпатии испытуемых. В подобных случаях положительный эффект сохранялся долго - до конца дня.
Дополнительные сенсорные нагрузки в виде быстрого увеличения освещенности визуального поля практически у всех испытуемых вызывали резкое ухудшение самочувствия, и в какой-то степени непроизвольные эмоциональные реакции, как правило, в виде произнесения междометий, ругательств, выражающих протест. Подобных реакций не было при аналогичных воздействиях при отсутствии у испытуемых симптомов дистресса. Длительное увеличение освещенности ухудшало проявления дистресса только у испытуемых, отличающихся сенсорной полезависимостью, и, напротив, могло уменьшать проявления дистресса у поленезависимых испытуемых.
Монотонные сенсорные нагрузки в виде световых мельканий, оптокинетических воздействий и монотонных акустических сигналов, как правило, усугубляли симптомы выраженного дистресса в условиях длительного вращения. Видимо, такие монотонные воздействия суммировались с монотонными гравитоинерционными воздействиями, т. е. с ведущим стрессором при жизнедеятельности в условиях длительного вращения, и это усиливало дистресс.
Исключением явились дополнительные строго ритмичные, дозированные по интенсивности гравитоинерционные нагрузки - они улучшали самочувствие испытуемых (исследование проводили Р. Р. Галле и Л. Н. Гаврилова). Причина их "исключительности", видимо, в следующем. Гравитоинерционные воздействия, возникающие при жизнедеятельности в условиях вращения, сложны и не упорядочены по своей ритмичности, интенсивности и направленности. Дополнительные "упорядоченные" гравитоинерционные стимулы, вероятно, в какой-то мере "упорядочивали" общий фронт гравитоинерционного стрессора, тем облегчая адаптацию к нему.
Вероятно, как усиление дистресса монотонии при дополнительном монотонном воздействии можно расценивать ухудшение самочувствия испытуемых в условиях вращения при прослушивании ими ритмичной рок-музыки. Данные, свидетельствующие об этом (совместно с Р. Р. Галле), следующие. Испытуемым предлагалось взять с собой любимые магнитофонные записи для прослушивания их в ходе многосуточного эксперимента с вращением (в "свободное время"). Регистрировалось, какие записи, сколько времени включали и слушали испытуемые. Было обнаружено, что в самом начале пребывания в условиях вращения, когда у ряда испытуемых возникала активизация субъективно приятных эмоционально-поведенческих реакций, проявлялся положительный эмоциогенный эффект музыкальных воздействий разного типа. В данном случае, можно полагать, суммировался субъективно-позитивный стрессогенный эффект гравитоинерционного и музыкального воздействий. Напомним, что такая "суммация" используется во многих увеселительных аттракционах в парках отдыха. Во второй фазе стресса, когда у испытуемых возникали проявления дистресса (чувство- слабости, головной боли, ощущение тяжести в животе, апатия, тошнота, рвота и т. д.), был отмечен различный эффект музыкальных воздействий разного типа. Практически у всех испытуемых при наличии у них выраженных проявлений дистресса рок-музыка и другая подобная монотонно-ритмичная музыка ухудшала самочувствие. Все испытуемые после нескольких проб отказывались от ее прослушивания. Некоторые любители такой музыки с удивлением сообщали, что неприязнь к ней сохранялась у них некоторое время после окончания многосуточного эксперимента с вращением. У большинства испытуемых при наличии у них симптомов выраженного дистресса было отмечено отчетливое предпочтение мелодичной музыки, а также эмоционально исполняемых песен с нетривиальным содержанием (например, записей песен в исполнении В. Высоцкого). По сообщениям испытуемых, субъективно неприятные симптомы дистресса в условиях вращения становились при этом менее заметными, улучшалось настроение.
Очевидно, что ритмичная рок-музыка создавала эмоционально-позитивные переживания за счет неспецифической эустрессовой мобилизации адаптационных резервов. Можно предположить, что при выраженном дистрессе монотонно-ритмичная музыка, "суммируясь" с повторяющимся гравитоинерционным стрессором, усугубляла симптомы дистресса. Мелодичная музыка и смысловое содержание песен могли снижать субъективно-неприятные переживания при дистрессе за счет отвлечения, а также за счет своей специфической адресации к позитивным эмоциям. Надо полагать, специальные исследования смогут обнаружить сложные психофизиологические и психологические механизмы, обусловившие отмеченные выше различия разных музыкальных воздействий при выраженном дистрессе.
Для иллюстрации вегетативных реакций при длительном, хроническом стрессе вновь обратимся к данным, полученным в 15-суточном эксперименте с непрерывным вращением испытуемых. В нижней части рис. 16 схематически представлены изменения самочувствия испытуемого Ко-ва. В ходе эксперимента оно значительно ухудшалось четыре раза: 1) при первом "столкновении" испытуемого со стрессором, 2) во время так называемой второй волны ухудшения самочувствия (см. ниже), 3) при действии дополнительного стрессора, возникшего из-за технической неисправности оборудования, и 4) сразу после остановки вращения, т. е. в начале реадаптации к условиям, лишенным стрессора (условий непрерывного вращения); в общих чертах такие изменения самочувствия были характерны для всех испытуемых, обследованных нами в аналогичных условиях. Кривая изменения содержания в крови испытуемого Ко-ва катехоламинов (регистрация Е. В. Елкиной) напоминает кривую мобилизации адаптационных резервов при стрессе по данным Г. Селье [241]. В первые часы действия стрессора содержание в крови катехоламинов уменьшалось. Можно предположить, что это результат их "убыточного" расходования. Вслед за этим уровень содержания катехоламинов в крови возрос существенно выше уровня, имевшегося до начала вращения, видимо, за счет мобилизации "глубоких" адаптационных резервов. Этот возросший уровень сохранялся до конца действия стрессора. Надо полагать, это отражало повышенное расходование адаптационных резервов как "оплату" организмом приобретенной резистентности (устойчивости) к стрессору. Содержание катехоламинов поддерживалось на высоком уровне, несмотря на значительное улучшение самочувствия испытуемого Ко-ва во второй половине эксперимента, с 8-х по 15-е сутки обитания во вращающемся помещении ("квартире"). Наряду с этим у испытуемого Ко-ва сохранялось малозаметное чувство неопределенного, трудно характеризуемого словами дискомфорта. В то же время в его поведении и высказываниях имели место постоянные попытки шутливо относиться к субъективно неприятным, тягостным симптомам кинетоза, сохранявшегося у него. Можно полагать, устойчивое чувство дискомфорта и высокий уровень катехоламинов крови - психологическое и физиологическое отражение адаптационных процессов, интенсивность которых устойчиво сохранялась, несмотря на смену за это же время ряда стадий в развитии стресса, отражавшихся в изменениях уровня других показателей стресса (см. рис. 16). Если резкий подъем содержания катехоламинов в крови наблюдался у всех людей в подобных условиях, то далеко не у всех этот уровень поддерживался в ходе эксперимента. При относительной свободе в выборе испытуемыми уровня двигательной активности содержание катехоламинов в крови значительно возрастало только в первые двое суток вращения до 23-28 мкг %, после чего уровень содержания в крови катехоламинов снижался и сохранялся умеренно повышенным до конца вращения и некоторое время после его прекращения. Подобный первоначальный подъем содержания в крови катехоламинов - показателя развития стресса - можно расценивать, во- первых, как избыточное, "на всякий случай" реагирование, во-вторых, как свидетельство "зашкаливания" интенсивности процессов адаптации выше "оптимального" уровня из-за недостаточности поначалу регулирующих ее механизмов, в том числе тормозящих двигательную активность и, таким образом, ограничивающих экстремальные воздействия. Содержание лейкоцитов в крови у Ко-ваг как и содержание катехоламинов крови, резко возросло в первые- вторые сутки вращения (регистрация Е. В. Елкиной). Однако в отличие от стабильно высокого уровня катехоламинов число лейкоцитов через двое суток действия стрессора начало снижаться и на 12-е сутки приблизилось к исходному, фоновому уровню. Видимо, на протяжении всего эксперимента сохранялась потребность организма в высоком содержании катехоламинов как в элементе "напряжения" организма, нужного для преодоления неблагоприятных эффектов стрессора. В отличие от этого первоначальный рост числа лейкоцитов в крови, т. е. превентивная защита от возможного проникновения в организм (и от возникновения в организме) чужеродного агента, оказался ненужным, так как такого агента не оказалось. Поэтому лейкоцитарная реакция, несмотря на продолжающееся действие гравиинерционного стрессора, угасла. Интересным является сопоставление показателей артериального давления в магистральных сосудах (в плечевой артерии), т. е. общего артериального давления, и локальных изменений артериального давления в центральной артерии сетчатки глаза, т. е. в ветви сосудов головного мозга (регистрация Б. В. Устюшина). Средний уровень общего давления крови у испытуемого Ко-ва несколько возрастал в ходе 15-суточного эксперимента с вращением (см. рис. 16). В этих условиях стали более выраженными суточные колебания артериального давления, что свидетельствует об увеличении лабильности (разбалансированности) вегетативной регуляции сердечно-сосудистой системы. Артериальное давление в русле мозговых сосудов в ходе эксперимента несколько раз резко возрастало. Каждый раз это сочеталось с ухудшением самочувствия испытуемого: у него в это время усиливались чувства слабости и дискомфорта, возникали апатия, головная боль, чувство тяжести в голове. Локальное повышение артериального давления в русле мозговых сосудов только по достижении определенного уровня сочеталось у всех обследованных испытуемых с ухудшением самочувствия, которое, в свою очередь, способствовало снижению психической и двигательной активности испытуемого. Это говорит в пользу "первичности" вегетативных реакций указанного рода и их "управляющего" влияния при стрессе на поведение и некоторые показатели психических процессов испытуемых. Приведенные данные являются примерами динамики при стрессе вегетативных функций трех типов: 1) "обслуживающих" развитие при стрессе психических феноменов, 2) "управляющих" этими феноменами, а также 3) "превентивно-защитных" в ситуации угрозы стрессогенного фактора. Первые сохраняют свой эффект, пока они необходимы. Вторые редуцируются в случае отсутствия агента, против которого они потенциально направлены. Третьи активируются, когда эмоциональная и поведенческая активность субъекта могут воспрепятствовать оптимально-экономному расходованию при стрессе адаптационных резервов. Вероятно, локальная спастическая реакция сосудов, сходная с обнаруженной в описанном выше эксперименте и в других подобных экспериментах, возникает при длительном "стрессе жизни". Допустимо предположить, что различные подобные локальные реакции оказываются предшественниками "болезней стресса": ишемической и гипертонической болезней, язвенной болезни желудочно-кишечного тракта и т. д. Именно подобные локальные вегетативные стрессогенные реакции могут явиться звеном в цепочке перехода стресса в болезнь. Общее (тотальное) повышение артериального давления, усиливающего кровоснабжение мозга и мышц, могло бы способствовать успеху агрессии или бегства. Локальное повышение кровяного давления только в сосудах слизистой оболочки желудка не способствует успеху защитного поведения. Более того, результатом таких локальных сосудо-суживающих реакций могут быть инсульт, инфаркт миокарда, точечные кровоизлияния в слизистой желудка и другие гибельные для субъекта (для особи) явления. Почему, вернее, зачем происходят эти зловещие проявления стресса? Для анализа причин этого феномена коротко повторим основные вехи нашего изложения отличий стресса у людей с активным и пассивным поведенческим реагированием в экстремальных условиях. При активном эмоционально-двигательном защитном (или агрессивном) стрессовом реагировании у субъекта формируется (неосознаваемо или осознанно) концептуальная модель развития ситуации с высокой вероятностью того, что стресс-фактор может быть удален прежде, чем исчерпаются адаптационные резервы организма. При пассивном ожидании того, что стрессор исчезает, за высокую вероятность принимается то, что уровень адаптационных резервов организма ниже необходимого для победы над стрессором. Поэтому принимается режим выжидания того момента, когда стрессор минимизируется. Это способствует экономии адаптационных ресурсов. Таким образом, достигается относительное "уменьшение" стрессора за счет адаптации к нему. При остром стрессе сохранение популяции возможно за счет особей как с активной, так и с пассивной поведенческой защитой от стрессора. Отбор продолжателей, носителей генофонда при этом происходит за счет губительного действия стрессора, т. е. за счет внешних по отношению к организму воздействий, которым организм не может противопоставить необходимую адаптацию или защиту. При длительном (прерывистом) стрессоре пассивное поведенческое реагирование и вместе с ним превентивно-защитная активизация вегетатики наблюдались практически у всех испытуемых. Такая форма защиты - более надежный путь к освобождению от стрессора, когда неизвестны или невозможны активные способы борьбы с ним. И тут оказывается, что индивиды, более склонные к пассивно-защитному поведению при сильном стрессоре (т. е. в некотором смысле менее целесообразно реагировавшие в таких условиях), оказываются быстрее приспосабливающимися к длительному (прерывистому) стрессору, болезненные проявления (симптомы) длительного дистресса у них хоть и более манифестированы, но менее субъективно неприятны, меньше снижают работоспособность.
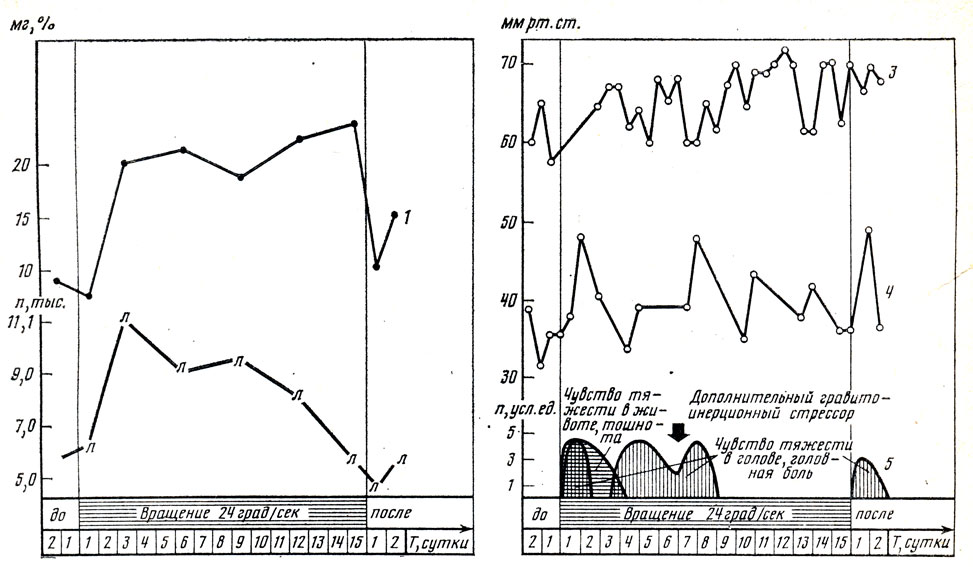
Рис. 16. Клинико-физиологические показатели при стрессе у испытуемого Ко-ва: 1 - катехоламины крови; 2 - лейкоциты крови; 3 - артериальное давление (минимальное) в плечевой артерии; 4 - в центральной артерии сетчатки глаза; 5 - показатели самочувствия
Можно предположить, что в данном случае проявляется один из основных законов биологического развития, закон экономии биологических ресурсов, лежащий в основе феномена индивидуальных различий живых организмов. В данном случае одни особи оказываются наделенными большей способностью активно, быстро преодолевать сильнодействующие экстремальные факторы, другие особи более успешны в пережидании неблагоприятных условий, которые могут затянуться. Остается неясным, почему, вернее, зачем для особей, менее успешно адаптирующихся при длительном стрессе, привентивно-защитные вегетативные реакции приобретают губительную "локальную" форму.
Прежде чем перейти к размышлениям над ответом на этот вопрос, нужно сделать следующее замечание. Как будет видно ниже, мы предполагаем, что в основе указанного проявления стресса лежат закономерности, выработанные в ходе биологической эволюции и целесообразные в животном мире. Однако эти эволюционные приобретения не могут быть полезны человеку ввиду отличий его социальной структуры жизни от жизни животных. Поэтому рассмотрим на биологических примерах основные вехи нашего изложения индивидуальных различий стресса (активное - пассивное защитное поведение; защитное поведение - вегетативная защита; тотальные защитные вегетативные реакции - локальное проявление таких реакций).
Согласно закону экономии биологических ресурсов особи с поведением, нецелесообразным для выживания популяции в условиях борьбы за существование, должны быть исключены из популяции, чтобы на них не расходовались экологические потенциалы, которые целесообразнее расходовать на особей, более успешных в адаптации к внешней среде, т. е. на более "полезных", "нужных" для сохранения, выживания популяции. Можно предположить два механизма, способствующих устранению менее адаптируемых особей из популяции при длительном, т. е. сравнительно несильном, не бьющем наповал, стрессоре. Это большая истощаемость адаптационных резервов у таких особей, т. е. меньший запас у них глубоких адаптационных резервов. Второй путь - самоуничтожение таких особей, происходящее по сигналу о неуспешности их адаптирования к требованиям среды и о необходимости сохранить экологический запас для расходования его членами популяции, более успешными в борьбе с окружающей средой. У нас нет данных для обсуждения первого гипотетического механизма устранения менее успешных особей. Вместе с тем есть примеры, говорящие о реализации в животном мире второго такого механизма. Приведем примеры.
У гусениц-шелкопрядов в определенном возрасте становится очевидным подразделение особей на более и менее жизнеспособных (больших и меньших по размеру). Менее жизнеспособные (до 50% популяций) падают с дерева и погибают. Остается столько особей, что им как раз хватает ресурсов питания, т. е. кроны дерева, на котором они живут [39, 2321. Такое уничтожение гусениц шелкопряда приходится оценивать как "беспричинную" гибель, когда квалифицированные анализы их трупов никаких признаков заболеваний не обнаруживают" [34, с. 431.
Другой пример - тотальная гибель поколения тихоокеанской горбуши раз в четыре года после окончания нереста. Разлагающиеся тушки погибшей рыбы служат кормом для молоди, которая, повзрослев, находит освободившиеся для нее от старшего поколения экологические пространства [79 и др.]. Следует отметить, что смерть горбуши происходит из-за инфаркта внутренних органов. Это указывает на то, что данный механизм используется в природе (отобран в ходе биологической эволюции) для как бы самоуничтожения особи, когда нет причин для ее смерти из-за неспособности противостоять болезни или от старости и т. д.
Допустимо предположить, что инфаркт, в частности, миокарда может служить не только для тотального уничтожения группы животных, сыгравших свою биологическую роль и поэтому лишних в популяции, но и для изъятия из популяции отдельных особей, оказавшихся "лишними" в популяции, т. е. по той или иной причине бесполезными или даже вредными для ее успешного существования. Возникает вопрос, как происходит "включение" подобного механизма "самоуничтожения" особи. Допустим такой схематический случай. Стая хищников гонится за жертвой, которая достается более быстрым и сильным особям. Их действия по достижению цели завершаются удовлетворением их желания, условно говоря, "торжеством победы". В данном случае осуществляется "комплекс активного реагирования" с его двумя фазами: фаза действия, фаза экстаза (см. выше). У особей, оказавшихся многократно (систематически) не успешными в достижении цели, будет накапливаться информация о систематическом отсутствии у них фазы "торжества победы". Как указывалось выше, это приводит особь к пассивному стрессовому поведению и к возникновению превентивно-защитных вегетативных реакций. Локальная модификация этих реакций может привести особь к гибели. В таком случае реализуется уже не защита особей от стрессора, а "защита" популяции от особи, входящей в ее состав, но не способствующей или препятствующей успешной жизнедеятельности и развитию популяции. Можно ли использовать результаты этих рассуждений для анализа "болезней стресса" в человеческом обществе? И до какой степени? Работами Фридмана и Розенмана было показано, что "человек типа А постоянно стремится выполнить слишком много или участвовать в слишком многих событиях за то количество времени, которое на это отведено... Поскольку он многократно создает такие непреодолимые преграды, он подвергает себя более или менее непрерывному давлению временем. Эта самотирания составляет суть поведения человека типа А. Наполнять жизнь непреодолимыми преградами и исключать все очарования жизни - это самая ужасная форма самонаказания" [384, с. 205]. К типу Б были отнесены люди противоположного склада. Люди типа А ("сизифов тип", "стресс - коронарный тип") значительно больше подвержены "болезням стресса". Они, в частности, в 6-8 раз чаще погибают от инфаркта миокарда, чем лица, относимые к типу Б [39].
Указанные примеры - иллюстрация фундаментальной диалектической закономерности, лежащей в основе индивидуальной разделенности живых существ, населяющих Землю, т. е. разделенности "биомассы", населяющей нашу планету, на особи. При этом смертность каждой особи призвана оптимизировать (ускорять) процесс адаптации популяции (вида, рода, биоценоза) к изменениям значимых для жизни физических факторов нашей планеты. "Смерть кажется жестокой победой рода над определенным индивидом и как будто противоречит их единству, - писал К. Маркс, - но определенный индивид есть лишь некое определенное родовое существо и как таковое смертен" [4, с. 119].
Современное индустриальное общество создает условия для дифференциации людей по их склонностям и проявлениям на людей типа А и типа Б. Человек часто оказывается в условиях, когда он еще до окончания одного задания знает о необходимости выполнения следующего задания и т. д. Спеша его выполнить, он лишает себя времени и условий для переживания радости по поводу окончания первого задания, а затем по поводу каждого очередного. Индивиды типа А более склонны активно входить в такой режим работы, режим жизнедеятельности, когда они оказываются систематически лишенными фазы "торжества победы". Вместе с тем люди этого типа больше нуждаются в ее переживании каждый раз после завершения фазы активной деятельности (преодоления трудностей, выполнения задания и т. п.) Лица типа Б менее склонны поддаваться давлению режима деятельности, лишающего их периодов позитивно-эмоционального переживания успеха окончания каждого цикла работы. Они менее чувствительны к такому лишению. В результате у индивидов типа А больше, чем у типа Б, накапливается информация об отсутствии должного быть после каждого завершенного действия, работы "торжества победы", т. е. якобы об их неуспешности в жизни и в деятельности. Парадоксальным является то, что не у представителей типа Б, а именно у людей типа А, часто более деятельных, энергичных, более успешных в делах, на которых поэтому больше "наваливается" работы и которые справляются с ней, в частности, за счет того, что отказывают себе в отдыхе, - именно у таких людей больше накапливается информации (может быть, неосознаваемой ими) об их якобы неуспешности. Так, надо полагать интерпретируется информация на биологических уровнях интеграции, информация о систематическом отсутствии "торжества победы" вопреки усилиям по ее достижению. Как правило, эта цепь событий не осмысливается субъектом. Однако, накапливаясь, подобная информация может вызвать у него чувство эмоционального дискомфорта, депрессивные проявления, может привести и к осознанию частой неуспешности своих действий, как причине дискомфорта.
Таким образом, целесообразный в животном мире механизм "защиты" популяции от ее членов, безуспешных в своих действиях, оказывается для человека нецелесообразным. Более того, этот механизм "популяционной селекции" в человеческом обществе во многих случаях оказывается ошибочно обратившимся против более успешных в своей деятельности и в этом смысле более полезных для общества людей.
Подытожим приведенную выше гипотезу. Действие длительного (многократного, монотонного) стрессора может вызывать у индивидов типа А, весьма способных к преодолению сильных кратковременных стрессоров, неблагоприятные для их организма вегетативные реакции. Эти реакции - локальные проявления вегетативной стрессовой активности (имеющей защитное значение в своем тотальном проявлении). Эти неблагоприятные, а часто губительные для организма реакции, надо полагать, возникли в ходе биологической эволюции животного мира. Они проявляются (реализуются) при длительных экстремальных ситуациях для "освобождения" популяции от особей, хронически неуспешных в своих действиях. Включаются эти губительные для особи (для индивида) локальные вегетативные реакции сигналами (вторичными) о накоплении сигналов (первичных) о регулярном отсутствии фазы "торжества победы", которая нормально должна завершать цикл деятельности (защитного) поведения (комплекс активного реагирования).
С точки зрения изложенной гипотезы могут быть объяснены некоторые неясные до настоящего времени симптомы стресса. Г. Селье пишет: "Механизм, посредством которого стресс вызывает язвы желудочно-кишечного тракта, также мало понятен". Мы предполагаем, что петехиальные кровоизлияния в слизистой оболочке желудка, впоследствии изъязвляющиеся, не имеют существенного защитно-адаптивного значения для особи (за исключением того, что их наличие ухудшает аппетит, что может быть иногда полезным при патологических и предпатологических состояниях). Вместе с тем эта патологическая стрессовая реакция в животном мире, видимо, целесообразна для "выбраковки" менее жизнеспособных членов популяции.
Указанные проявления дистресса возникают вследствие локальных сосудистых спазмов, приводящих к локальным дистрофиям слизистой оболочки желудка и двенадцатиперстной кишки, которые усугубляются локальной экскреторной реакцией (только в желудке или только в двенадцатиперстной кишке). Неблагоприятную для особи роль выполняют при стрессе и другие локальные проявления защитных вегетативных реакций: локальный спазм сосудов внутренних органов (может привести к инфаркту сердечной мышцы, легких, печени, селезенки и т. д.), локальная гиперплазия ткани (доброкачественная или злокачественная опухоль) и т. д.
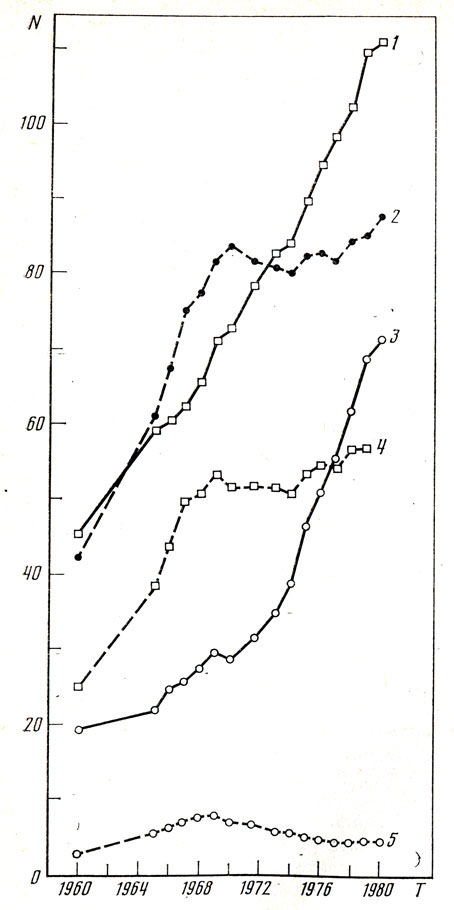
Рис. 17. Смертность населения СССР от сердечно-сосудистых заболеваний (на 100000 человек): 1 - сосудистые поражения головного мозга (без гипертонической болезни), 2 - гипертоническая болезнь, 3 - ишемическая болезнь сердца (без гипертонической болезни), 4 - сосудистые поражения мозга при гипертонической болезни, 5 - инфаркт миокарда при гипертонической болезни (Вестник статистики, 1969, № 2; 1970, № 6; 1971, № 2, № 12; 1973, № 12; 1975, № 12; 1977, № 12; 1979, № 11; 1981, № 11)
Клинические наблюдения указывают на то, что во многих случаях (далеко не всегда) психосоматические больные гипертонической болезнью не заболевают язвенной болезнью желудка, а "язвенники" отличаются невысоким артериальным давлением. Э. Гельгорн и Дж. Луфборроу, указывающие на эту закономерность, отмечают, что она связана с "генетической конституцией", люди первого типа - симпатикотоники, второго типа - ваготоники [61, с. 374]. Является ли столь разноплановая дифференциация больных на "гипертоников" и "язвенников" проявлением какого-то единого функционального процесса? Напрашивается аналогия указанной дифференциации и деления людей: одних - склонных к тотальным, других - к локальным вегетативным проявлениям дистресса. Ответ на такой вопрос - дело будущего. Между тем ответ на него чрезвычайно важен в связи с ростом "болезней стресса", среди которых одни протекают по типу тотальных реакций, другие проявляются в виде локальных поражений. Среди болезней сердечно-сосудистой системы выделяют гипертоническую болезнь (протекающую на фоне тотальной реакции сердечно-сосудистой системы) и такие виды патологии как протекающие без гипертонической болезни ишемическую болезнь сердца и поражение сосудов головного мозга (преимущественно локальные проявления болезни). На рис. 17 и табл. 2 видны различия в смертности от этих заболеваний. Обращает внимание, что ишемическая болезнь сердца, часто афишируемая как наиболее распространенная среди "болезней стресса", не является таковой в ряде стран (Япония, СССР и др.), где первое место среди причин смертности занимает поражение сосудов мозга. Следует шире смотреть на часто приводимые сведения о большей заболеваемости мужчин, чем женщин, ишемической болезнью сердца [60, 155 и др.]. Это действительно так. Вместе с тем нельзя не учитывать, что смертность от значительно более распространенных поражений сосудов головного мозга мало отличается у мужчин и у женщин (рис. 18).
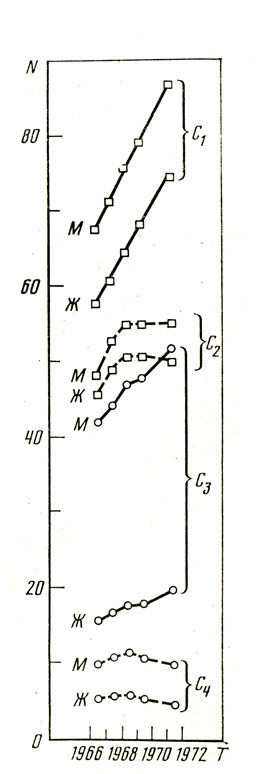
Рис. 18. Смертность населения СССР от сердечно-сосудистых заболеваний с разделением по полу (на 100 ООО человек) (стандартизированные данные): М - мужчины, Ж - женщины. C1 - сосудистые поражения головного мозга (без гипертонической болезни) (М/Ж-1,16); С2 - сосудистые поражения головного мозга при гипертонической болезни) (М/Ж-1,1); С3 - ишемическая болезнь сердца (без гипертонической болезни) (М/Ж-1,26) С4 - инфаркт миокарда при гипертонической болезни (М/Ж-2,1). (Вестник статистики, 1969, № 2; 1970, № 6; 1971, № 2; 1971, № 12; 1973, № 12)
Можно подразделять локальные стрессовые вегетативные реакции на экскреторные (гиперацидная секреция слизистой желудка и т. п.), эргатические (спазм сосудов и т. п.), дистрофические (экзема, атрофия десен и т. п.), воспалительные (наружные, локализующиеся в кожных покровах, и внутренние, во внутриорганизменных органах и тканях) и т. д. Подробному описанию таких реакций посвящена обширная медицинская литература. К сожалению, ее авторы часто игнорируют анализ роли психологических факторов в возникновении указанных выше локальных вегетативных патогенных реакций. Не учитывается психическое состояние, имевшееся перед началом болезненных реакций и в ходе заболевания. Такой недоучет психических стрессогенных, патогенных факторов уменьшает "прицельность" и эффективность лечебных мероприятий. Учитывать роль психики в развитии болезней, в том числе тех, которые отнесены сейчас в разряд "болезней стресса", призывали многие выдающиеся представители медицины: Пирогов, Захарьин, Боткин, Ланг, Бурденко, Мясников и др.
Предлагаемая нами гипотеза пока имеет мало обосновывающих и подтверждающих ее фактов. Тем не менее ввиду исключительной актуальности проблемы "болезней стресса" для населения индустриальных стран, ввиду безотлагательности предотвращения этих болезней мы считаем целесообразным использование данной концепции при разработке мероприятий по борьбе с неблагоприятными проявлениями стресса.
Основные принципы борьбы с "болезнями стресса" согласно предлагаемой концепции состоят в следующем. 1) Проводить анализ жизнедеятельности населения и трудовых процессов с выделением их естественной микро- и макроцикличности. При отсутствии "естественного" структурирования таковой цикличности создавать ее "искусственно". Обеспечивать эмоционально-усиленное переживание субъектом успешности и полезности завершения каждого трудового цикла, каждого цикла жизнедеятельности. 2) В тех жизненных, производственных условиях, где индустриализация лишает человека чувства успешности преодоления трудностей и опасностей, необходимо периодически моделировать условия, порождающие такие чувства. Такое моделирование необходимо избирательно для субъектов, склонных к преодолению трудных или опасных ситуаций. 3) Разрабатывать и внедрять методы рекреакции, направленные на восстановление способности к оптимальной актуализации "комплекса активного реагирования" (с его основными двумя фазами) прежде всего у людей, утративших такую способность. При организации такой рекреации необходимо учитывать личностные и психофизиологические особенности людей, а также социокультурные и этнические особенности их жизни.
О субъективной вероятности, субъективной "невозможности" и экстремальности измерения среды. Для удобства анализа описанных выше явлений представляется целесообразным формулирование ряда определений некоторых особенностей среды с позиции адаптивной, биологической системы, подвергающейся воздействиям со стороны среды.
![Таблица 2. Показатели смертности в некоторых капиталистических странах в 1955-1973 гг. (на 100 000 человек) [28]](pic/000019.jpg)
Таблица 2. Показатели смертности в некоторых капиталистических странах в 1955-1973 гг. (на 100 000 человек) [28]
В ходе фило- и онтогенетического развития биологическая система на основании прецедентов общения (столкновения) со средой (внешней и внутренней) накапливает в своей памяти (памяти в широком смысле слова) фонд "комплексов" адаптивного реагирования. Каждый из них, условно говоря, имеет в своем составе предназначенную для сличения с реальным событием концептуальную модель значимого для данной системы изменения среды (события, ситуации), программу реагирования, адекватного отраженному в концептуальной модели изменению внешней среды, и аппарат функционально-структурного обеспечения этого комплекса реагирования. Говоря о памяти в широком смысле слова, мы имеем в виду запечатленный в структурно-функциональных особенностях биологической системы и в ее специфическом аппарате памяти результат отражения по ходу ее развития, динамики ее "общения" со средой. Значимое изменение среды, значимое событие - это событие, могущее изменить гомеостатическое состояние биологической системы. Минимальной значимостью изменения среды определяется возникновение порога абсолютной чувствительности биологической системы к данному изменению среды. Значимое событие, вызывающее изменение гомеостаза биологической системы, угрожающее нарушением или нарушающее ее целостность, жизнеспособность, - это опасное (экстремальное) событие (изменение среды). Чувствительность биологической системы к минимально опасному изменению внешней среды является порогом чувствительности к опасности (опасному событию).
Изменения адаптивной системы в ответ на изменения внешней среды (экзогенное побуждение) в соответствии с внутренними программами адаптации (эндогенным побуждением), т. е. реагирование системы, - это проявление ее активности в широком смысле.
Активным в узком смысле реагированием биологической системы будем называть ее реагирование, направленное на ликвидацию опасности среды путем ликвидации носителя опасности или увеличением расстояния между ним и реагирующей системой (отбрасывание, изгнание носителя опасности или уход от него). Ниже мы будем использовать термин "активное реагирование" в узком смысле. Реагирование адаптивной системы в ответ на опасное изменение среды (на носителя опасности) путем ожидания либо исчезновения опасности, либо завершения адаптивной перестройки (в соответствии с эндогенными программами адаптации), в результате которой опасный фактор, даже не изменившись, перестанет быть опасным для данной биосистемы, данный тип реагирования мы называем пассивным реагированием.
То или иное изменение среды, значимое или опасное для биологической системы, может быть в разной мере вероятным. Субъективная вероятность события обусловлена числом его прецедентов в ходе развития этой системы. Субъективная вероятность значимого (опасного) события прямо пропорциональна активности ответного реагирования системы. Чем более многократно встречалась биологическая система в ходе своего развития с аналогичными событиями, тем больше ее адаптивные возможности (активность разного рода) для сохранения ее гомеостатического равновесия (целостности, жизнестойкости) вопреки влиянию среды. В ходе развития биологической системы возникали те или иные значимые и опасные для нее изменения среды, в то же время каких-то (какого-то вида) изменений среды не возникало. И это не могло не отразиться в памяти (в широком смысле) данной биологической системы. В структурно-функциональной организации достаточно сложной целостной системы в ходе ее развития сформировались программы реагирования, предназначенные для предотвращения гомеостатических нарушений при хоть сколько-нибудь вероятных значимых событиях. Иными словами, функционально-структурный состав адаптивной системы, его логика, его закономерности определены кругом значимых событий той или иной вероятности. Беспрецедентные события, даже если они гипотетически значимы или опасны для адаптивной системы, не могут учитываться (со всей их спецификой) закономерностями, логикой построения этой системы. Подобные беспрецедентные для данной системы изменения среды (события, ситуации) "невозможны" (назовем их так) для данной системы. В теории вероятностей существует определение "невероятного" события как пустого подмножества во множестве всех подмножеств пространства элементарных событий данного опыта. "Невероятное" событие "не происходит ни при каком исходе опыта" [245, с. 180]. В данной интерпретации "невероятные" события равны нулю и недифференцированы.
Представляется целесообразным иное понимание термина "невозможное" событие. Следует предположить, что в ходе развития рассматриваемой биологической системы имели место "невозможные" события разного вида. После каждого такого события последующие события этого же вида воспринимались этой системой как события "возможные", т. е. в той или иной степени вероятные. События этого вида находили свое отражение в соответствующих адаптационных перестройках биологической системы. Надо предположить, что биологическая система должна была отразить в своем строении саму возможность возникновения класса "невозможных" событий, т. е. сформировать какие-то формы реагирования для защиты от гипотетической опасности "невозможных" событий. При невозможности реагировать на значимое возникновение "невозможного" события из-за отсутствия программ адекватного, в достаточной мере специфического реагирования биологическая система должна иметь программы неспецифических форм реагирования, адекватных всему классу "невозможных" изменений среды. При этом допустимо предположить возникновение в ходе развития адаптивной биологической системы способности оценивать "степень невозможности" беспрецедентного события. Эта, так сказать, выраженность невозможности, классифицируемая биологической системой, должна зависеть от способности этой системы определять степень несоответствия вновь возникшей (новой) "невозможной" ситуации той логике, закономерностям строения данной биологической системы, которые сформированы в процессе ее развития, т. е. при ее столкновении с "невозможными" событиями иного вида, чем происшедшее в текущий момент.
Изменения среды могут быть более или менее информативны для биологической системы в плане определения ею возможности или невозможности этого изменения среды. Изменение более информативное в этом смысле можно назвать более определенным. Можно говорить об определенности события достаточной или недостаточной для установления (идентификации) биосистемой "возможности" или "невозможности" данного события и о степени (о выраженности) "возможности" (вероятности) или "невозможности" этого события.
Субъективная оценка изменений среды биологической системой (организмом) обусловливает те или иные особенности реагирования последней. Незначимые (подпороговые) изменения среды не "включают" какого-либо значимого реагирования системы, но могут воздействовать на нее на подпороговом уровне (т. е. критерии значимости условны!). Значимая, но неопределенная информация об изменения среды обусловливает ориентировочное реагирование (поведение) организма, направленное на увеличение определенности события. При достаточной определенности изменений среды "включаются" программы специфического целенаправленного реагирования биологической системы. Информация об опасных изменениях внешней среды "включает" программу защитного активного или пассивного реагирования. Наконец, есть основания выделить некоторого рода комплексные изменения внешней и внутренней среды организма, которые могут породить сигналы (управляющую информацию) этому организму о целесообразности его реагирования, например о целесообразности его "самоликвидации", в целях сохранения или повышения жизнеспособности не данного организма, а популяции, в которую он входит. Естественно, в реальных условиях существования в рамках высокоорганизованного организма (особи, биологической системы) вместе с "включением" механизмов его самоликвидации начинают действовать защитные для данной особи механизмы, направленные, во-первых, против (на нейтрализацию) процессов "самоликвидации", во-вторых, на исчерпание ситуации, при которой целостность особи противопоставляется "жизнеспособности" популяции. Мы остановимся на этом ниже.
Следует отметить еще, что разные по иерархии уровни биологической системы в рамках организма особи специализированы на разных видах, родах активности, в частности защитной активности. Высоковероятные опасные изменения среды "включают" активное эмоционально-двигательное защитное реагирование, вегетативная сфера имеет при этом "обслуживающее" вторично-защитное значение. При "невозможных" событиях "включается" пассивное эмоционально-двигательное реагирование; значительная активация при этом вегетативной сферы приобретает первичнозащитное значение.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://psychologylib.ru/ 'Библиотека по психологии'