57. Исследование сенсорной настройки как психофизиологического выражения целевой установки методом регистрации вызванных потенциалов. Л. А. Самойловым, В. Д. Труш ( A Study of Sensory Attunedness as Psychophysiological Expression of Goal-Directed Set by the Method of Recording Evoked Potentials L. A. Samoilovich, V. D. Trush
57. Исследование сенсорной настройки как психофизиологического выражения целевой установки методом регистрации вызванных потенциалов. Л. А. Самойловым, В. Д. Труш
МГУ, факультет психологии
Одно из направлений в области исследования психологии установки было связано в последнее десятилетие с изучением процессов селекции во время переработки сенсорной информации. Основная дискуссия в рам.ках этого направления развернулась вокруг проблем, связанных с локализацией механизмов селекции: одними исследованиями показывалось, что селекция возникает в момент акта восприятия - гипотеза входа [10], в других работах отмечалась возможность фильтрации на выходе системы - гипотеза ответа [16]. В то же время результаты ряда работ позволяют предположить, что избирательность существует на протяжении всего процесса переработки информации, т. е. селективное преднастроечное влияние задачи простирается на все уровни акта восприятия [2; 9; 11; 14; 16].
Для объяснения психофизиологических механизмов, реализующих установки в деятельности, неоднократно привлекались представления о доминанте А. А. Ухтомского [7], образе потребного будущего Н. А. Бернштейна [3], акцепторе действия П. К. Анохина [1], нервной модели стимула Е. Н. Соколова [6]. Однако конкретно-экспериментальное исследование психофизиологических механизмов, реализующих процессы селекции, стало возможным лишь с появлением моделей, описывающих процессы переработки информации как микроструктуру, которая состоит из последовательно включающихся функциональных блоков [4]. В рамках таких моделей анализ мозговых механизмов селекции в условиях установки может быть выполнен с помощью исследований электрических явлений коры головного мозга. Одним из широко используемых методов анализа электрических корковых явлений является регистрация вызванных потенциалов (ВП), которые развиваются после предъявления любого сенсорного сигнала и представляют собой последовательность волн, создаваемых активностью различных мозговых систем [5; 8]. Связи ВП с процессами внимания посвящен обширный литературный материал, однако до сих пор в электрофизиологической литературе дискуссируется вопрос о том, в какой степени наблюдаемые изменения в различных компонентах ВП связаны с процессами внимания [15].
Настоящее исследование было направлено на выяснение связи ВП зрительной коры мозга человека с явлениями перцептивной селекции, создаваемой разными целевыми установками.
Экспериментальная часть работы была выполнена на основе микроструктурной модели процессов восприятия, в условиях быстрой предъявления последовательности сенсорных сигналов. Исследование состояло из двух экспериментов: в первом эксперименте введение установки не приводило к успешной селекции тестового материала, во втором - процесс селекции полностью снимал маскировку тестовых стимулов.
Исследование проведено на 5 испытуемых с нормальным зрением. Для предъявления материала использовался экспериментальный стенд, состоявший из люминесцентного индикатора и пульта ответов, соединенных с ЭВМ. В каждом эксперименте было проведено по 5 серий: контрольная без установки и четыре серии с установкой "на знакоместо". Во всех сериях предъявлялось по 100 проб, каждая из которых начиналась с нажатия испытуемым кнопки "готовности", после чего через 500 мсек вспыхивала цифра "0" (сигнал "внимание"). Через 1 сек после этого сигнала предъявлялась стимульная последовательность, состоявшая из 4 цифр с временем свечения каждой цифры 20 мсек и межстимульным интервалом 80 мсек. Цифры з стимул ьно и последовательности выбирались ЭВМ случайным образом из алфавита "2-9" таким образом, чтобы ни одна из цифр не встречалась в последовательности дважды. Через 1 сек после предъявления последней цифры в последовательности испытуемому предъявлялась послестимульная цифра-инструкция, выбираемая ЭВМ из того же алфавита таким образом, чтобы вероятность ее появления в последовательности составляла 0,6.
В контрольных сериях испытуемый опознавал цифровой материал на всех четырех знакоместах; перед началом тестовых серий он получал установочную инструкцию - опознать цифру только на определенном знакоместе (в первой серии на I знакоместе и т. д.). Цифры на остальных знакоместах выполняли при этом функцию маски. Второй эксперимент отличался от первого лишь тем, что в тестовых сериях в качестве маски использовалась фиксированная цифра (например - "5"), а не весь алфавит "2-9".
В ходе всего опыта регистрировалась электроэнцефалограмма (ЭЭП, которая отводилась монополярно от затылочных областей коры головного мозга. Усреднение вызванных потенциалов производилось после опыта на ЭВМ, программа которой позволяла производить синхронное усреднение одновременно с подсчетом текущих значений дисперсий. Усреднение производилось для каждой серии по 100 ответам, после чего полученные кривые ВП и их дисперсий выводились на двухканальный графопостроитель.
Хорошо выраженный "v"-образный характер позиционных кривых в контрольных сериях указывает на наличие наибольшей маскировки средних элементов в последовательности, вероятность правильных ответов на которые падает до 0,57 (±0,08) для S2 и до 0,65 (±0,07) для S3 (рис. 1А). Анализ позиционных кривых, построенных по результатам тестовых серий первого эксперимента показал, что в условиях установки на знакоместо маскировка снимается лишь для крайних знакомест S1 и S4. Замена маски, состоящей из полного алфавита (2-9), на маску в виде фиксированной цифры во втором эксперименте оказала существенное влияние на успешность опознания средних элементов последовательности. Таким образом, во втором эксперименте оказалась возможной эффективная селекция знакоместа, что приводило к успешному опознанию тестового материала на любом знакоместе, снимая маскировку всех элементов последовательности. Улучшение опознания отражалось и на позиционных кривых времени реакции испытуемых (рис. 1Б).
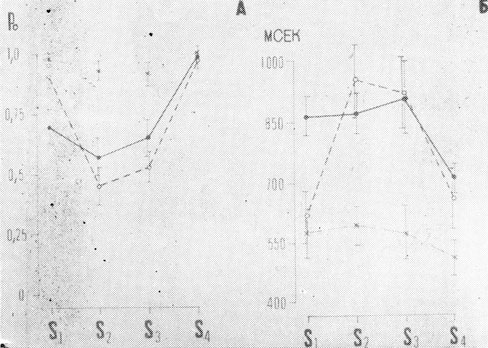
Рис. 1. Позиционные кривые вероятности правильных ответов (А) и зависимость времени реакции для правильных ответов от знакоместа стимула в последовательности (Б). (По оси ординат порядковые номера знакомест. Сплошная линия - контрольные условия, пунктирная - эксперимент I, штрих - пунктир - эксперимент II. Вертикальными линиями на кривых обозначен доверительный интервал при р=0,01, критерий Стьюдента)
У всех испытуемых на одиночный сенсорный сигнал (Mi) наиболее выраженными были компоненты ВП с пиковыми латентностями: для первого положительного компонента 100+10 мсек, первого отрицательного 140±20 мсек и второго положительного 210±40 мсек (рис. 2). В связи с тем, что определение изолинии в ходе обработки ВП оказалось не всегда возможным, амплитудные значения компонентов ВП определялись как разница максимальных значений двух соседних пиков, то есть анализировалась амплитуда P1-N1 и N1-P2.
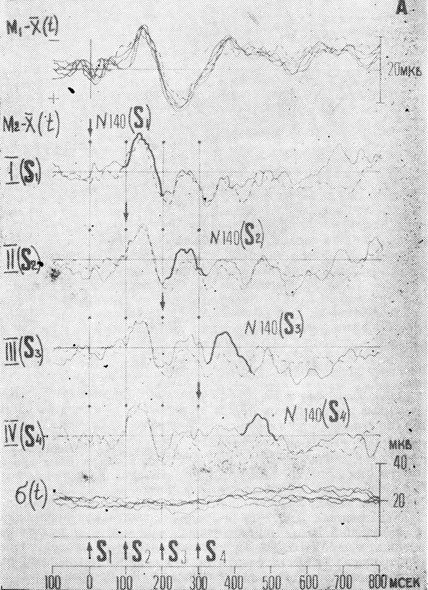
Рис. 2. Вызванные потенциалы, зарегистрированные в зрительной коре в разных экспериментальных условиях. Пунктирная линия - ВП в контрольных условиях, сплошная - ВП в условиях установки при отсутствии селекции (А) и при ее наличии (Б). (М1 - одиночный ВП, М2 - ВП в условиях маскировки, σ - суперпозиция кривых среднеквадратичных отклонений для всех условий. Вертикальными римскими цифрами и стрелками указывается, для какого знакоместа в данной серии осуществляла сенсорная настройка)
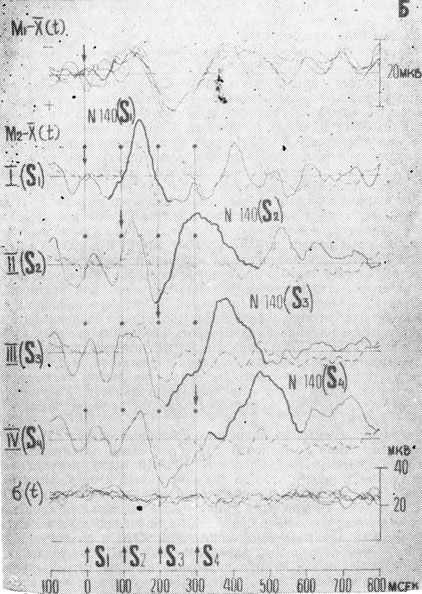
Рис. 2. Вызванные потенциалы, зарегистрированные в зрительной коре в разных экспериментальных условиях. Пунктирная линия - ВП в контрольных условиях, сплошная - ВП в условиях установки при отсутствии селекции (А) и при ее наличии (Б). (М1 - одиночный ВП, М2 - ВП в условиях маскировки, σ - суперпозиция кривых среднеквадратичных отклонений для всех условий. Вертикальными римскими цифрами и стрелками указывается, для какого знакоместа в данной серии осуществляла сенсорная настройка)
Прежде чем исследовать характеристики ВП в условиях перцептивной селекции, были проанализированы амплитудно-временные изменения компонентов ВП в контрольных условиях при последовательной зрительной маскировке (рис. 2, пунктирные кривые). Как видно из рисунка, ответы на второй, третий и четвертый стимулы в последовательности существенно уменьшаются. Наиболее сильное подавление амплитуды при маскировке наблюдалось для компонентов N1-Р2 (с 23 мкв для S1 она снижалась до 7-9 мкв для S3 и S4; р=0,01, критерий Стьюдента). Рис. 3. Для перехода Р1-N1, уменьшение амплитуды также носило статистически достоверный характер (с 14 мкв до 6-7 мкв, р=0,01). Полного восстановления анализируемых компонентов не наблюдалось даже для стимула S4, который следовал через 300 мсек после первого стимула в последовательности, когда развитие ответа на одиночный стимул практически заканчивается. Одновременно с уменьшением амплитуды компонентов ВП при маскировке наблюдалось статистически значимое удлинение латентных периодов компонентов ВП на маскируемые стимулы (в среднем на 40 мсек дли S2 и на 30-40 мсек для S3 и S4, рис. 4).
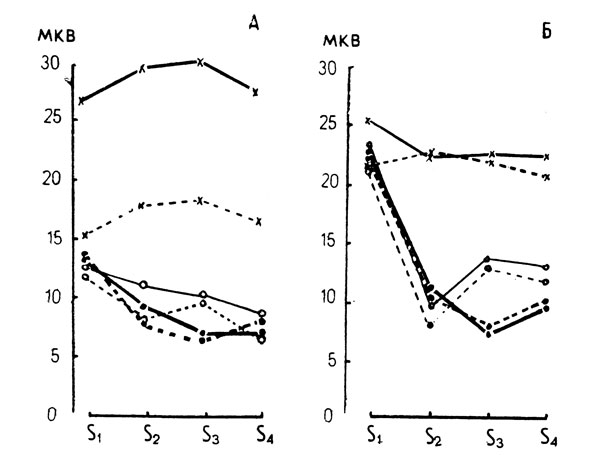
Рис. 3. Изменение амплитуды компонентов Р100-N140 (А) и N140-Р200 (Б) в условиях целевой установки на различное знакоместо. (Сплошная линия - правое полушарие, пунктир - левое, ● - контрольная серия, ○ - эксперимент I, Х - эксперимент II)
В вызванных потенциалах, зарегистрированных в первом эксперименте (тестовые серии с установкой на знакоместо при отсутствии селекции), не было обнаружено статистически значимых изменений (р=0,5) для всех анализируемых компонентов ВП (рис. 2, 3, 4). Хорошо выраженное влияние установки на ВП наблюдалось во втором эксперименте в условиях селекции. Амплитуда компонентов P1-N1 при селекции возрастала в 1,5-2 раза для всех знакомест. Амплитуда волны P1-N1 при успешной селекции максируемых знакомест S2, S3 восстанавливалась по сравнению с контрольными условиями при маскировке и была равной величине ответа на S1 (р=0,01). Повышение эффективности процессов селекции приводило к сильному сдвигу латентных периодов ВП, регистрируемых на средние элементы последовательности с 140 мсек для S1 до 220-240 мсек для S2 и S3. На стимул S4 латентность ВП уменьшалась до величины латентности в контрольных условиях (рис. 4).
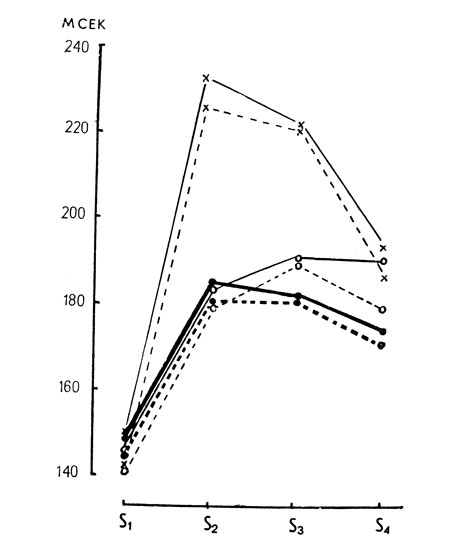
Рис. 4. Изменения латентности компонента 140 в условиях целевой установки на знакоместо (обозначения в подписи к рис. 3)
Установленная в экспериментах связь процессов селекции с увеличением амплитуды компонента N140 зрительного вызванного потенциала позволяет предположить существование психофизиологического механизма селекции на сенсорно-перцептивном уровне. Доказательством того, что процесс селекции в наших исследованиях протекает на довербальном уровне, являются данные по природе и функциональному значению регистрируемых компонентов ВП [5, 8].
Полученные данные хорошо согласуются с результатами исследований ВП в условиях селективного слухового внимания [12; 13]. На основе этих исследований Хиллярд предположил существование двух различных механизмов избирательного внимания: первый обеспечивает избирательное пропускание сенсорного сигнала в определенном канале, а второй - выбирает конкретный перцептивный признак внутри ранее выделенного канала и связан с вербально-категориальным уревнем анализа. Усиление N140 в этих экспериментах Хиллярд связывает с "поддержанием тонической установки" предпочтения определенного входа, изменения в Р300 связываются им с процессами сличения сенсорного образа с заготовленными эталонами.
В заключение необходимо отметить, что целевые установки не только меняют процесс восприятия, но и, как показывают результаты настоящего исследования, активно формируют мозговые функциональные системы, реализующие эти процессы.
Литература
1. Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968.
2. Асмолов А. Г., Ковальчук М. А., Яглом М. А., Об иерархической структуре установки. В сб.: Новое в психологии, 1975, 1, 10-24.
3. Бернштеин Н. А., О построении движений, М., 1947.
4. Зинченко В. П., О микроструктурном методе исследования познавательной деятельности. В сб.: Эргономика. Труды ВНИИТЭ, вып. 3, М., 1972.
5. Иваницкий А. М., Мозговые механизмы оценки сигналов, М., 1976.
6. Соколов Е. Н., Механизмы памяти, М., 1969.
7. Ухтомский А. А., Очерк физиологии нервной системы, Л., 1954.
8. Шага. С. Ч., Вызванные потенциалы в норме и патологии, М., 1975.
9. Bachmann, Т.. Allik. I.. Integration and interruption in the masking of form by form. Perception, 1976, v. 5, 79-97.
10. Erdelyi, M. L., A new look at the New Look: Perceptual defease and vigilance. Psychological Review. 1974, v. 81, 1-25.
11. Haber. R.N., Nature of the effect of set on perception. Psychological Review. 1966. v. 73. 335-351.
12. Hillyard. S.A., Hink. R. F., Schwent. V. L. and Picton. T.W.. Electrical signs of selective attention in the human brain. Science, 1973, 182, 177-180.
13. Hillyard S. A., Schwent, V. L., Evoked potential correlates of selective attention with nulti-channel auditory inputs. EEG and Clin. Neurophysiology, 1975. 38. 131-138.
14. Krueger. L. E.. Familiarity effects in visual information processing. Psychological Bulletin, 1977, v. 82. 949-974.
15. Naatanen. R., Selective attention and evoked potentials. Ann. Acad. Sci. Fenn. В., 1977, 151, 1-226.
16. Shiffrin. R.M., The locus and role of attention in memory systems. In: Rabbit, P.M., Dornic. S. (Eds.) Attention and Performance. V. S. Academic Press, 1975.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://psychologylib.ru/ 'Библиотека по психологии'