6.2. Поведение при стрессе в эволюционном аспекте
Данные, приведенные в предыдущем разделе, свидетельствующие в пользу наличия генетической обусловленности индивидуальных различий поведения при стрессе, были получены на грызунах, т. е. на биологическом объекте, весьма отдаленном в эволюционном плане от человека. Интересным было проследить характер стрессовых реакций у животных, стоящих на разных уровнях эволюционного развития.
Ниже приведены результаты исследования поведенческих реакций различных животных при возникновении у них стресса во время многократных повторений кратковременной невесомости (табл. 5) [115, 118].
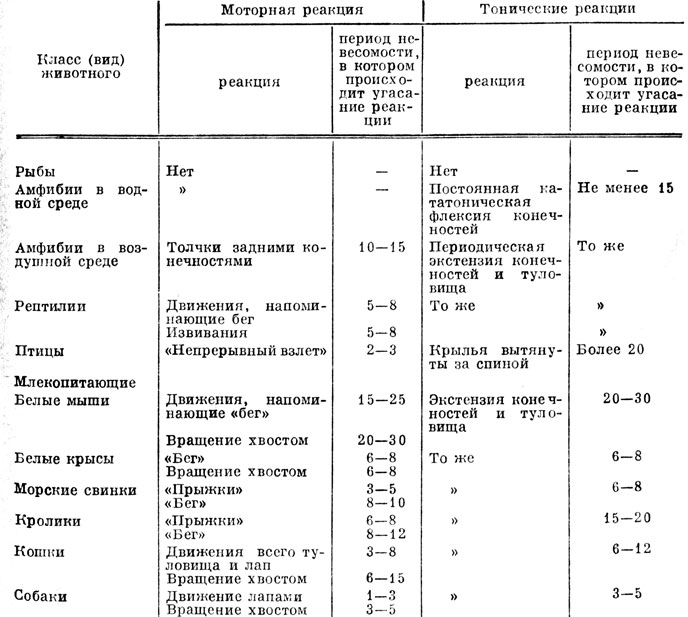
Таблица 5. Продолжительность адаптации к многократному действию невесомости у различных животных
В экспериментах были исследованы поведенческие реакции животных, принадлежащих разным уровням эволюционного развития: рыб (золотых рыбок), земноводных (лягушек), рептилий (ящериц), птиц (голубей), млекопитающих (белых мышей, белых крыс, морских свинок, кроликов, кошек и собак). Использовали по 10 животных каждого вида без учета возраста и пола. В кабине самолета были оборудованы специальные отсеки, установлены контейнеры, в которых животные могли свободно парить при невесомости. В экспериментах с рыбами контейнеры заполнялись водой. Лягушек исследовали при невесомости в водной и в воздушной средах. Поведение животных регистрировалось с помощью киносъемки с последующей покадровой расшифровкой. В разных режимах полета у животных определялись число и характер движений, а также определялось положение животных относительно "верха" и "низа" кабины самолета.
У рыб в невесомости существенного изменения интенсивности движений не отмечено. Вместе с тем в тех случаях, когда рыба, плывущая вперед внезапно оказывалась в невесомости (в режимах полета, когда невесомость создавалась сразу вслед за состоянием с естественной силой тяжести, т. е. без перегрузки, предшествовавшей невесомости), было замечено, что с исчезновением силы тяжести рыба как бы замирала. Замирание с подрагиванием плавниками было также характерным для рыб при перегрузке.
В невесомости у рыб утрачивалась ориентация относительно верха и низа аквариума. При этом рыбы могли оказаться в положениях - брюхом "вверх", боком "вверх", "вниз" головой и т. п. (относительно координат аквариума). Только после окончания режима невесомости они мгновенно принимали положение спиной вверх. В нескольких случаях было отмечено, что при повторных пребываниях в невесомости рыбы за счет движений плавниками и хвостом (а не за счет перемещений вместе с массой воды) начинали переворот вокруг горизонтальной оси: головная часть продвигалась "вниз", хвостовая - "вверх" (относительно координат туловища рыбы). При этом из положения "спиной вверх" рыба перемещалась в положение "хвостом вверх" и т. д. Ввиду технического несовершенства методики не удалось обнаружить изменений объема туловища рыб при невесомости и при перегрузке, что могло бы свидетельствовать об их реакциях, направленных либо на всплытие, либо на погружение.
Особого внимания заслуживает различие двигательной активности земноводных в воздушной и иммерсионной средах. В первом случае у всех лягушек возникало бурное усиление движений в виде чередования выпрямления и сгибания конечностей (1-5 циклов в секунду). Движения могли прекратиться только в случае, если животное фиксировалось в банке, либо упираясь головой и конечностями в противоположные стенки банки (на это бы ли способны крупные животные), либо "прилипнув" к стенке банки. Следовательно, прекращению активизации движений при невесомости в воздушной среде способствовало возникновение тактильной или тактильно-кинестетической информации о наличии опоры, т. е. о в некотором роде стабильности окружающего пространства. У всех лягушек в водной среде во время невесомости возникала пассивная двигательная реакция. У них возникала стабильная поза, характеризующаяся напряженным сгибанием конечностей, известная под названием "поза молельщика".
Рептилии во время парения в невесомости совершали частые движения лапами и движения туловищем в виде "извивания".
У птиц после исчезновения действия силы тяжести возникали маховые движения крыльями в виде непрерывного взлета, в результате они летели "вверх", к потолку, переворачивались и летели к полу, далее снова к потолку и т. д. о второго, третьего режима (см. табл. 5) эта реакция угасала и птицы повисали в воздухе в своеобразной позе с крыльями, вытянутыми за спиной, и с растопыренными перьями хвоста; они могли цепляться за обшивку кабины, сохраняя позу с вытянутыми крыльями. Начиная с 20-го режима птицы зависали в невесомости со сложенными крыльями и хвостом.
Двигательная активность млекопитающих в невесомости усиливалась. Первоначально возникали движения, напоминающие бег животного: у мышей и крыс - "барабанящие", реципрокные движения выпрямленными конечностями, у всех прочих - "прыжки" (чередующиеся сгибания и разгибания туловища) и "барабанящие" движения конечностями. Одновременно у млекопитающих, кроме морских свинок и кроликов, возникали вращательные движения выпрямленным хвостом; в результате реактивного момента туловище начинало вращаться в противоположную сторону. Повышение двигательной активности всегда сопровождалось возбуждением животных: криком, расширением глазных щелей, у кошек - выраженной хватательной реакцией. Все эти реакции с самого начала невесомости протекали на фоне выраженной экстензии конечностей и запрокидывания головы назад. В экспериментах отмечена активизация некоторых вегетативных функций млекопитающих: усиление саливации, мочеиспускания и дефекации (которые начинались в невесомости), у кошек перспирация. Случаев рвоты не было. По мере возникновения адаптации к невесомости, при многократном ее повторении (см. табл. 5), первоначально исчезали "прыжки", затем "барабанящие" движения лапами. Животные переставали вращать хвостом, продолжая держать его при невесомости вытянутым в дорзальном направлении. После исчезновения двигательных реакций экстензия конечностей и туловища становилась более заметной. По мере адаптации животных экстензорная реакция исчезала. При отсутствии силы тяжести они повисали, замерев в воздухе, поджав лапы к животу, выпрямляя их только, чтобы схватить за какой-либо предмет, оказавшийся поблизости. Поведение кошек и собак отличалось по сравнению с прочими животными более быстрым наступлением адаптации к невесомости и большим индивидуальным разнообразием реакций.
Нельзя утверждать, что невесомость совершенно незнакома земным существам. Исчезновение силы тяжести сравнительно часто испытывают различные организмы в условиях наземного существования. Возникая лишь при свободном падении, невесомость, по-видимому, сигнализирует об опасности возможного удара о землю и не длится, как правило, более 2-3 сек. Нам представляется, что механизмы физиологических реакций, вызванных исчезновением силы тяжести, следует рассматривать, исходя из представлений о динамике процессов, которые возникают в организме при свободном падении. Известно, что у животных падение вызывает экстензорную (лифтную) реакцию конечностей и туловища, вращательные движения хвостом (они стабилизируют животное в потоке встречного воздуха), активизацию хватательных рефлексов. Биологическое значение этих моторных актов - в подготовке позы, обеспечивающей наиболее безопасное приземление.
Подобные реакции отмечены [310, 146 и др.] у животных и в начальном периоде невесомости в полетах па самолетах и спутниках. В двигательпом возбуждении можно выделить [115, 118] тоническую (экстензорную) реакцию конечностей и туловища и моторную реакцию - хватательные движения, вращательные движения хвостом, реципрокные движения конечностей в виде прыжков и бега. Сходство моторики в начальном периоде невесомости и при падении свидетельствует о том, что этот период, по-видимому, воспринимается животными как падение. Невесомость, продолжающаяся более 2-3 сек., не имеет каких-либо фило- или онтогенетических прецедентов, и, становясь сигналом чрезвычайной опасности, вызывает интенсивную оборонительную реакцию "убегания" (движения в виде прыжков и бега, крик, расширение глазных щелей и пр.).
Г. Мэгун [202] указывает, что реципрокное сгибание конечностей, сочетающееся с прогибанием спины, постоянно возникает при прямом раздражении ретикулярной формации, причем невозможность вызвать ретикулоспинальное торможение объясняется функциями коры, снижающей возбудимость продолговатого мозга. Удаление областей вестибулярного представительства в коре препятствует развитию двигательного возбуждения в невесомости у кошек. Чрезмерная выраженность реципрокных движений может расцениваться как результат растормаживания подкорки: реципрокные движения конечностей характерны для эмбрионального периода до сформирования функций коры.
При невесомости, создаваемой в закрытых кабинах самолетов, спутников и т. п., информация, поступающая от гравирецепторов (вестибулярного, кожно-мышечного) и ряда интероцепторов, свидетельствует об исчезновении реакции опоры, о падении, т. е. об опасном для организма животного или человека нарушении стабильности окружающего. В то же время зрение сигнализирует об отсутствии пространственных изменений, вокруг видны те же стены, пол и потолок кабины. Таким образом, возникают два противоречивых потока информации. Анализируя работу различных сенсорных систем, Хольст [421] предположил, что в случае, когда от разных рецепторов поступают противоречивые сообщения, в низших мозговых центрах происходит "нейтрализация", "гашение" их и высших центров достигает только "остаток преобладающего сигнала".
Результаты экспериментов с животными, полученные в условиях кратковременной невесомости, подтверждают высказанные выше суждения. Очевидно, в силу преобладающего значения в пространственном восприятии животных вестибулярного и двигательного анализаторов по сравнению со зрительным [12, 32], после исчезновения силы тяжести у животных возникали моторные реакции, характерные для свободного падения в естественных условиях, в соответствии с гравирецепторной информацией. По мере наступления адаптации к условиям невесомости моторная активность животных снижалась.
Продолжительность адаптирования к повторяющимся воздействиям невесомости, можно полагать, зависит не столько от эволюционного уровня животного, сколько от его экологических особенностей. Рыбы и земноводные, обитая в иммерсионной среде, практически лишены реакции опоры. Вместе с тем они могут перемещаться в трех измерениях, что создает естественную адаптацию к небольшим изменениям силы тяжести. Можно полагать, все это обусловило отсутствие у них в невесомости двигательного возбуждения. О значении тактильных сигналов в формировании поведения в невесомости говорит значительное различие характера подвижности земноводных при невесомости в водной и в воздушной среде. Относительно короткий период адаптирования птиц, вероятно, тоже вызван их естественной адаптацией к уменьшениям и увеличениям силы тяжести, часто возникающим в полете. Период адаптации к невесомости у кошек и собак короче, чем у мышей и кроликов, вероятно, потому, что зрительный анализатор у хищников более высоко развит, чем у грызунов.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://psychologylib.ru/ 'Библиотека по психологии'