Эмоция
"Процессы установки стереотипа, довершения установки, поддержки стереотипа и нарушений его и есть субъективно разнообразные положительные и отрицательные чувства".
"В каждом знании столько истины, сколько математики".
Существует притча о двух лягушках, попавших в банку со сметаной. Одна из лягушек, убедившись, что выбраться из банки невозможно, прекратила сопротивление и погибла. Другая продолжала биться, хотя ее прыжки и казались бессмысленными. Под ударами лягушачьих лапок сметана постепенно загустела, превратилась в комок твердого масла. Лягушка влезла на него и... выпрыгнула из банки.
Эта маленькая сказка хорошо иллюстрирует две самые характерные черты эмоционального напряжения. Эмоция возникает при недостатке сведений, необходимых для достижения цели. Замещая, компенсируя этот недостаток, она обеспечивает продолжение действий, способствует поиску новой информации и тем самым повышает надежность живой системы.
К решению вопроса о приспособительной роли эмоций ближе всех подошел И. П. Павлов. Он создал учение о динамическом стереотипе - устойчивой системе ответных реакций животного и человека, соответствующей определенной комбинации внешних сигналов, Было замечено, что осуществление даже очень сложного, но знакомого стереотипа действий не сопровождается теми характерными сдвигами дыхания, сердечной деятельности, кровенаполнения сосудов, которые позволяют говорить об эмоциональном напряжении. Более того, напряжение не возникало и при некоторых изменениях системы внешних сигналов, если только эти изменения не вели к переделке стереотипа реакций. Но как только внешний стереотип условных сигналов начинал требовать изменения ответных действий, появлялись признаки эмоции. Животное с выработанным динамическим стереотипом представляет систему, располагающую полной информацией о том, когда, что и как следует делать. Нарушение стереотипа означает появление дефицита информации, необходимость поиска сведений для организации новой деятельности по удовлетворению потребности в пище, избегании болевого воздействия и т. д.
Эмоция возникает каждый раз, когда удовлетворения потребности не происходит, иными словами, когда действия не достигают цели. Если у человека выработать условный оборонительный рефлекс таким образом, что исследуемый в ответ на условный раздражитель имеет возможность отрывать пальцы от электродов, то изменения сердечного ритма исчезают по мере выработки устойчивой двигательной реакции. Если же электроды одеты на пальцы и оборонительное движение руки не избавляет от действия тока, условные сердечные рефлексы оказываются стойкими и значительными по величине (Л. Я. Балонов, 1959).
"Ну и что же, - скажет внимательный читатель, - все дело сводится к раздражению током. Если человек получает удары тока, он волнуется, его сердце бьется учащенно. Если, отрывая пальцы, он не получает тока, сердце успокаивается". Действительно, в нашем примере человек с кольцами на пальцах наверняка знает, что тока ему не избежать. Но такова природа эмоционального аппарата: организм не хочет примириться с неизбежностью боли, он бьет тревогу учащенными ударами сердца и требует поиска выхода из любого положения. Опыт с кольцами на пальцах поставлен противоестественно. Человек по договоренности с экспериментатором сознательно подавляет оборонительную реакцию и не срывает кольца, на которые подается ток. Изменим опыт таким образом, чтобы он больше походил на естественное поведение в оборонительной обстановке.
Человеку дается задание: нажимать на кнопку через 20 секунд после звукового сигнала. Если он ошибается больше, чем на 1 секунду (до или после условленного момента), на кожу руки подается болезненный ток. При попадании в "цель", т. е. в интервал от 19 до 21 секунды, тока не бывает. После каждой реакции исследуемому сообщают ее истинное время, чтобы он мог исправлять допущенные ошибки. Рис. 1 и 2 показывают результаты опытов на двух исследуемых. Сплошная кривая линия отмечает время двигательной реакции, причем полоса в нижней части рисунка обозначает "зону безопасности", при попадании в которую человек избавляет себя от боли. Верхняя пунктирная кривая показывает частоту сердцебиений - суммарную протяженность на записи электрокардиограммы трех первых ударов сердца после звукового сигнала и трех последних перед двигательной реакцией. Тонкой горизонтальной линией обозначен исходный фон сердечной деятельности - средняя протяженность записи шести сердечных сокращений. Благодаря этой линии мы можем судить о степени учащения сердечного ритма по сравнению с состоянием покоя.
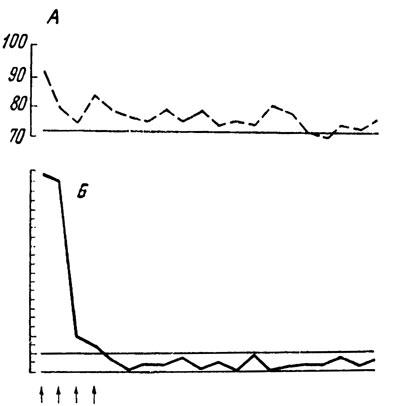
Рис. 1. Опыт с выработкой условного оборонительного рефлекса на время: А - изменения частоты сердцебиений (число сокращений в минуту); Б - время двигательных реакций в секундах. Стрелки обозначают раздражения электрическим током
Рассматривая рис. 1, мы видим, как у человека постепенно вырабатывается условный двигательный оборонительный рефлекс на время. После первого периода грубых ошибок исследуемый "нащупывает зону безопасности" и начинает попадать в цель. Почти одновременно исчезает состояние эмоционального напряжения - ритм сердцебиений возвращается к исходному уровню. При таком ходе опыта может создаться впечатление, что эмоция тревоги, страха, о которой мы судим по учащению сердечного ритма, целиком зависит от болевых раздражений: есть удары тока - ритм учащен, прекратились раздражения током - человек успокоился, сердце бьется ровно. Вот почему для нас гораздо важнее те случаи, где человеку далеко не сразу удается выработать защитный рефлекс (рис. 2). При сравнении участка опыта от 2-го до 15-го применения условного сигнала с участком от 16-го до 29-го применения гудка мы видим, что человек ошибся и, следовательно, получил ток одинаковое число раз (по шесть ударов). Если бы эмоция зависела только от болевых раздражений, суммарное учащение сердцебиений на первом и втором этапах опыта (показано штриховкой) должно было бы составить равную величину. Однако этого но произошло. На первом этапе сердце билось сильнее, чем на втором. Чему же соответствует учащение сердцебиений при равном количестве болевых раздражений строго постоянной силы? Суммарному отклонению двигательных реакций от цели (показано черным цветом), т. е. недостатку сведений о том, каким именно должно быть движение, избавляющее человека от боли! Аналогичные отношения наблюдаются и при сравнении проб 30-38 с пробами 39-47.
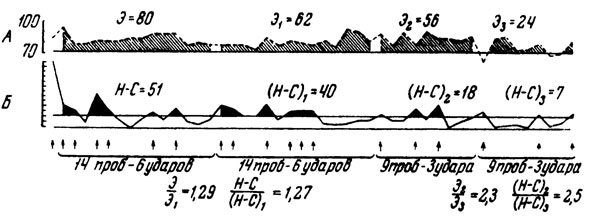
Рис. 2. Сопоставление суммарного учащения сердцебиений по сравнению с фоном (А) и суммарного отклонения времени двигательных реакций от заданной величины (Б) на разных этапах опыта. Стрелки обозначают раздражения электрическим током. Можно видеть, что степень эмоционального напряжения (Э), о котором мы судили но уменьшению сердцебиений, изменяется пропорционально с уменьшением суммарного отклонения от цели Н - С)
В общем виде это правило можно выразить формулой:
Э = П (Н - С),
где Э - эмоция; П - потребность (побуждение); Н - информация, прогностически необходимая для организации действий по удовлетворению данной потребности; С - информация, которая может быть использована для целенаправленного поведения.
Из данного выражения с неизбежностью вытекают минимум четыре следствия:
1. Э = 0 при П = 0; эмоция не возникает в отсутствие потребности и исчезает при ее удовлетворении.
2. Э = 0 при Н = С; эмоция не возникает (или резко ослаблена) у вполне информированной системы даже при больших значениях П. Эмоцию нельзя связывать только с фактом потребности (ее семантикой: голодом, жаждой, опасностью и т. д.), она имеет прагматическую природу и существует в системе "потребность - действие - удовлетворение". Ярчайший пример вполне информированной системы - павловский динамический стереотип, реализация которого почти не сопровождается признаками эмоционального напряжения.
3. Э максимальна, если С = 0; при наличии цели (потребности) живая система в известных пределах тем эмоциональнее, чем менее она информирована.
4. При С больше Н Э изменяет свой знак. Поскольку в приведенной формуле мы рассматриваем потребность (П) как побуждение, отрицательное в биологическом смысле для организма, случай С больше Н должен вести к возникновению положительной эмоции. Этот случай будет подробно проанализирован несколько позднее.
Формулу эмоций можно проверить, подставив в нее результаты опыта с рефлексом на время. Эмоция будет выражаться числом сердцебиений, потребность - напряжением электрического тока, а разность (Я - С) - величиной ошибки, допущенной исследуемым при осуществлении двигательной реакции. Конечно, эта проверка носит относительный характер "первого приближения" к действительности. Ведь эмоция характеризуется весьма разнообразными сдвигами в организме, а отнюдь не только изменением частоты сердцебиений. Да и оценка потребности в устранении боли по величине электрического тока тоже условна. Тем не менее экспериментальная проверка формулы дает вполне удовлетворительные результаты.
Нужно ли говорить, что еще большие трудности возникают при попытке количественно оценить степень эмоционального напряжения в случаях, когда мы имеем дело с эмоциями высшего порядка, с потребностями, сформированными условиями воспитания, с информацией, необходимой для осуществления сложных действий человека. В этих случаях, пожалуй, наиболее трудно измерить именно потребность, и здесь формула может оказаться очень полезной. Если мы замерим эмоцию по величине физиологических сдвигов в организме, а для оценки дефицита сведений воспользуемся достижениями теории информации, то величину потребности можно будет рассчитать математически. Ведь, согласно формуле, П будет равно Э/(Н - С).
Количественные расчеты сложных эмоциональных состояний в настоящее время трудно осуществимы. Пока что формула имеет преимущественно теоретическое значение, позволяет кратко выразить самое главное в природе эмоций - их зависимость от потребности и дефицита прагматической информации.
С развиваемой точки зрения эмоция представляет компенсаторный механизм, восполняющий дефицит информации, необходимой для достижения цели (удовлетворения потребности). Термин "информация" мы всюду употребляем с учетом ее содержательной ценности. В этом смысле возрастание количества информации означает повышение вероятности достижения цели благодаря получению данной суммы сведений. Можно возразить, что накопление сведений, необходимых для достижения той или иной цели, происходит в процессе познания. Но, говоря об эмоции, мы имеем в виду не прирост сведений (он возникает в процессе поиска, движущую силу которого составляет потребность, усиленная эмоциональным напряжением), а именно компенсацию, замещение, экстренный механизм, включающийся на данном этапе познания при данной степени информированности о возможных путях удовлетворения возникшей потребности.
Мы полагаем, что приведенное определение эмоции обладает тремя положительными свойствами: а) отражает физиологическую сущность явления, б) показывает связь эмоции с потребностью и действием, равно как и различие между ними, в) оказывается универсальным для любого из известных нам эмоциональных состояний. Так, ярость компенсирует недостаток сведений, необходимых для организации борьбы (известно, что исчерпывающие сведения о наилучшем поражении противника делают борьбу "хладнокровной"). Страх возникает при недостатке информации, требующейся для организации успешной защиты. Отсюда поведение, инициированное страхом, это - бегство или неподвижность (ступор), биологическую целесообразность которого специально подчеркивал И. П. Павлов. Испуг (его нельзя смешивать со страхом, потому что страх возникает до угрожающего воздействия, а испуг следует за воздействием) - внезапный дефицит сведений об источнике и размерах неожиданной угрозы. Горе мы наблюдаем в ситуации острейшей недостаточности информации о какой бы то ни было возможности восполнения потери (ср. утешение).
Наконец, повседневный опыт показывает, что мы прибегаем к эмоциональным средствам воздействия (например, к подчеркнуто эмоциональной речи) чаще всего при известной нехватке строго логических доводов или в обстановке, когда для изложения доводов нет времени. С этой точки зрения повышенная эмоциональность детей и подростков объясняется не "свежестью чувств" или какими-то особыми свойствами молодого мозга, а недостатком жизненного опыта, хронической "недоинформированностью" относительно происходящих вокруг событий и возможности удовлетворения своих формирующихся потребностей. Положение о "сдерживающем" влиянии коры па подкорковые образования мозга и "возбуждающем" влиянии подкорки, повторенное в сотнях статей, нуждается в объяснении. Почему кора преимущественно тормозит нервный аппарат эмоций? Почему эмоциональные центры преимущественно возбуждают высшие отделы мозга? В каких случаях кора оказывает сдерживающее, а в каких - стимулирующее влияние на подкорковые центры? Это становится понятным, если учесть, что кора представляет у высших животных и человека основное "хранилище" прагматического опыта, а центры эмоций сосредоточены в подкорковых отделах.
Можно привести множество примеров компенсаторного значения эмоций. Если потребность в достижении цели достаточно велика, ярость помогает человеку бороться в тех условиях, где его поражение кажется совершенно очевидным. Чувство тревоги удерживает от опрометчивых поступков при самых отдаленных и слабых признаках опасности. Воодушевление, трудовой энтузиазм позволяют достигнуть результатов; которых не в состоянии предвидеть трезвый расчет. "Мы можем ровно столько, сколько знаем", - писал в свое время Ф. Бекон. По-видимому, его определение следует дополнить: мы можем столько, сколько знаем, плюс то, что дает нам эмоциональное напряжение. В отдельных случаях логический расчет, ослабляя степень эмоциональной мобилизации, способен затруднить достижение цели. Об этом очень ярко написал Сент-Экзюпери, рассказывая о летчике, который потерпел аварию в безлюдных горах. "...Начиная со второго дня, больше всего уходило сил на то, чтобы не думать..., положение было уж очень безнадежным, чтобы иметь мужество идти, надо было поменьше размышлять о своем положении"*.
* (Л. де Сент-Экзюпери. Земля людей. М., 1957, стр. 104.)
Особенно велика роль эмоции как движущей силы поиска информации, недостающей для удовлетворения потребности. Сколько люден проявили невероятное упорство, разгадывая тайны природы и общества. Подчас они тратили на это всю свою жизнь, хотя вероятность достижения цели была исчезающе мала, "...без "человеческих эмоций" никогда не бывало, нет и быть не может человеческого искания истины"*.
* (В. И. Ленин. Сочинения. Изд. 4-е, т. 20, 1948, стр. 237.)
До сих мор мы говорили главным образом об эмоциях, которые принято называть отрицательными. В какой мере наше определение справедливо для положительных эмоций? Напомним, что знак эмоции изменяется в случае, когда имеющаяся информация превышает прогностически необходимую. Действительно, каждый раз, когда возникает положительное эмоциональное состояние, живая система оказывается избыточно информированной, причем степень положительного переживания тем больше, чем значительнее разница между действительностью и прогнозом.
...Томимый жаждой путник движется по раскаленным пескам. Он знает, что только через три дня пути сможет встретиться с источником. Удастся ли пройти этот путь? Хватит ли сил? Не занесло ли ручей песками? И вдруг, завернув за выступ скалы, человек видит колодец, не отмеченный на картах. Бурная радость охватывает усталого путника. В тот момент, когда перед ним блеснуло зеркало колодца, путник стал обладателем исчерпывающих сведений о возможности утоления жажды. И это в ситуации, когда прогноз в лучшем случае предсказывал три дня тяжелейших скитаний! Разумеется, сила положительной эмоции (как и отрицательной) зависит от величины потребности. Вторым определяющим фактором является величина (Н - С) - контраст между прогнозом и реальной действительностью.
Человек бесстрашен, когда он обладает исчерпывающими сведениями о том, как выйти из опасного положения. Опыт, мастерство, совершенное владение техникой - вот основа бесстрашия летчика-испытателя, верхолаза, циркового артиста. Нетрудно видеть разницу между "безумством храбрых", действующих наперекор недостатку информации о возможности достижения цели (об этом мы будем подробно говорить в следующей главе), и бесстрашием человека, обладающего избыточной информацией. В первом случае страх был подавлен другой, более мощной потребностью, во втором - избыток сведений разрушил страх "изнутри", превратил эмоцию страха в свою противоположность.
Чувство торжества возникает только у того, кто преодолел большие трудности на пути к достижению цели. Чем смелее была поставлена задача, чем недостижимее казалась цель, иными словами, чем значительнее разрыв между достигнутым и первоначально имевшимся в распоряжении, тем сильнее торжество победителя. Это свойство положительных эмоций очень точно подметила норвежская писательница Сигрид Унсет: "Хорошие дни выпадают на долю разумных людей, но лучшие дни достаются тому, кто посмеет быть безумным".
Каково приспособительное значение положительных эмоций? В чем заключается их компенсаторная роль? Когда мы говорим о достижении цели, мы всегда берем какой-то отрезок, фрагмент приспособительного поведения. Ведь маршрут путешественника, о котором шла речь выше, не окончился возле счастливо встреченного колодца. Тонизирующее влияние положительных эмоций необходимо для дальнейшего движения вперед, для достижения новых целей. Руководители экспедиций, рабочих коллективов, воинских групп хорошо знают, как важен бывает хотя бы небольшой первоначальный успех для дальнейшей деятельности. Успех ободряет, порождает чувство уверенности в своих силах. Компенсируя дефицит информации о путях достижения цели, положительная эмоция побуждает человека ставить эту цель и действовать в намеченном направлении. Это свойство положительных эмоций нельзя абсолютизировать. Торжество победителя может обернуться "головокружением от успехов", утратой ориентировки в реальной действительности и заслуженным поражением. Но в известных пределах именно положительные эмоции обеспечивают быстрое продвижение вперед за счет "перешагивания" через "информационные пустоты". Вероятностно изменяющаяся среда коварна и многолика. Самый опытный летчик не застрахован от встречи с неведомой опасностью. Но когда он встречается с этой опасностью, чувство уверенности, сформированное предшествующим опытом, не допускает возникновения страха, способного помешать поиску выхода из положения.
Таким образом, положительные эмоции радости, торжества, бесстрашия в полной мере обладают компенсаторными свойствами, но, в отличие от отрицательных эмоций, эти свойства обнаруживаются не в отдельном приспособительном акте, а в целостном процессе адаптивного поведения.
Какие нервные механизмы лежат в основе эмоционального реагирования? Какие особенности этих механизмов обеспечивают ту роль, которая принадлежит эмоциям в сложной деятельности живых систем? Мы но будем перечислять физиологических теорий, имеющих в настоящий момент чисто историческое значение, и остановимся только на современных исследованиях. Центральный нервный аппарат эмоций представлен совокупностью мозговых образований, которые принято называть "висцеральным мозгом" (Пейпец, 1037, 1962; Клювер и Бьюси, 1938; Мак-Лиан, 1953; Гельгорп. 1961). Название подчеркивает тесную связь всех образований "висцерального мозга" с регуляцией деятельности внутренних органов. Эта связь имеет двоякий смысл. С одной стороны, она напоминает о непосредственной зависимости эмоций от органических потребностей организма (голод, жажда, половое влечение и т. д.); с другой - делает попятными те глубокие сдвиги вегетативных функций, которые паблгодаются при реализации эмоции в сердечно-сосудистой системе, дыхании, деятельности желез внутренней секреции, органах пищеварения и выделения. К "висцеральному мозгу" относятся: подбугорная область (гипоталамус), передние ядра таламуса, перегородка, свод мозга, маммилярные тела, миндалевидное ядро и образования древней коры (венечная борозда, гиппокамп, грушевидная доля). "Висцеральный мозг" тесно связан с новой корой больших полушарий, особенно с ее лобными, височными и теменными долями.
Функциональные влияния центрального аппарата эмоций можно разделить на два основных типа: восходящее и нисходящее. Восходящее влияние состоит главным образом в активации, возбуждении, повышении работоспособности высших отделов мозга и органов чувств. Осуществляя это влияние, "висцеральный мозг" тесно взаимодействует с ретикулярной формацией. Дело в том, что сама ретикулярная формация, активизирующее значение которой стало очевидно после исследований Моруцци и Мэгупа (1949), физиологически пассивна и нейтральна в эмоциональном отношении (Кардо, 1961). Она вовлекается в деятельность регулирующими командами с коры больтних полушарий (Жуве, 1956, 1961; С. Н. Нарикашвили, 1961; Моруцци, 1962; А. И. Ройтбак и С. М. Бутхузи, 1961, 1963 и мн. др.), а также импульсацией из центров эмоций, прежде всего из гипоталамуса. Стимуляция заднего гипоталамуса усиливает вызванные корковые ответы па свет и звук (Гельгорн, 1961а), т. е. даст эффект, типичный для ретикулярной активации. Разрушение гипоталамуса существенно изменяет характер влияния ретикулярной формации па высшие отделы мозга. Если в нормальных условиях достаточно сильные периферические раздражении будят спящее животное и десинхронизируют его электроэнцефалограмму (ЭЭГ), то после разрушепия задпего гипоталамуса активация ЭЭГ ие сопровождается пробуждением (Фельдман и Воллер, 1962). Можно думать, что досинхронизация ЭЭГ преимущественно связана с холинэргической системой, центры которой представлены в переднем отделе гипоталамуса, а реакция пробуждения - с адренэргической системой заднего отдела. Известно, что введение ацетилхолина активирует ЭЭГ, не изменяя поведения животного, в то время как адрепаловая активация сопровождается истинным пробуждением (О. С. Виноградова, 1961).
Существует мнение, что именно гипоталамусом задаются основные ритмы электроэнцефалограммы (Генри и Сковилл, 1952). По свидетельству ряда авторов, эмоциональное возбуждение отражается на электроэнцефалограмме в виде реакции активации: подавление альфа-ритма наряду с усилением десинхронизированной активности. Линдсли считает эту картину типичной для состояний настороженности и боязливости (1960, 1961). Блокада альфа-ритма при счете в уме усиливается, если счет приобретает эмоциональную окраску, благодаря вознаграждению за правильный результат (Глесс, 1964). Наши собственные наблюдения, а также данные литературы дают основание считать, что депрессия альфа-ритма наиболее характерна для ориентировочной реакции. Если фактор, вызывающий эмоциональное напряжение, утрачивает свою новизну, депрессия альфа-ритма отсутствует даже при сильных эмоциях, сопровождающихся значительными вегетативными сдвигами в организме.
Хотя возбуждение ретикулярной формации возникает вторично, оно существенно влияет на гипоталамические эффекты. После разрушения ретикулярной формации среднего мозга раздражения шейных симпатических нервов, заднего гипоталамуса и введение адреналина не вызывают реакции активации в коре больших полушарий (А. М. Мариц, 1963). Стимуляция переднего отдела ретикулярной формации ствола усиливает, а продолговатого мозга - усиливает и тормозит кожногальванический рефлекс у кошек. Центральное представительство этого рефлекса находится в гипоталамусе (Вонг, 1956; Покота Тошикацу, 1963).
Весьма своеобразные отношения существуют между ретикулярной формацией и таким важным звеном эмоционального аппарата, как гиппокамп. При воздействиях, вызывающих в коре больших полушарий десинхронизацию электрической активности (вспышки света, раздражение гипоталамуса), в гиппокампе регистрируются большие медленные волны с частотой 5-7 колебаний в секунду (Грин и Ардуини, 1954). Аналогичный эффект наблюдается при введении кролику эзерина (Бремер, 1961). Весьма вероятно, что так называемый ритм напряжения с частотой 4-7 в секунду, зарегистрированный в электроэнцефалограмме животных и человека при отрицательных эмоциях (Г. Уолтер, 1953; П. К. Анохин, 1963а), имеет гиппокампальпое происхождение. Знаменательно, что ритм 3-5 колебаний в секунду, возникающий в гиппокампе при раздражении седалищного нерва, после разрушения медиального гипоталамуса исчезает и сменяется десинхронизацией (Коразза и Пармеджиани, 1963). Это наблюдение указывает на причастность гипоталамических структур к особенностям биоэлектрической активности гиппокампа. Эксперименты дают основание говорить о тормозящем влиянии гиппокампа на ретикулярную формацию мозгового ствола (Лишак и Граштьян, 1962).
Таким образом, имеющиеся в настоящее время данные свидетельствуют о том, что восходящие влияния физиологического аппарата эмоций в значительной мере реализуются через сетевидное образование. Это, конечно, не исключает прямого действия эмоциональных центров на кору больших полушарий, а также влиянии через симпатический и парасимпатический отделы вегетативной нервной системы, столь обстоятельно изученного выдающимся советским физиологом Л. А. Орбели.
Обширные нисходящие эффекты эмоционального возбуждения реализуются через вегетативную нервную систему, высшие центры которой локализованы в гипоталамусе. При этом активность парасимпатического отдела преимущественно связана с передним и латеральным гипоталамусом, а симпатического отдела - с задним и медиальным (Гельгорн, 1961; Линдсли, 1960 и др.). Бовард (1961, 1962) предложил гипотезу о существовании в мозгу высших животных и человека двух функционально противоположных систем. Одна из них имеет парасимпатическую природу и представляет физиологическую основу положительных эмоций. Вторая, адренэргическая система включается при отрицательных эмоциях. Раздражение нерпой из этих систем вызывает чувство удовольствия VI служит наградой для подопытного животного. Пациенты с электродами, вживленными в центры позитивной системы, нередко просят врача о повторных раздражениях. Стимуляция негативной системы сопровождается беспокойством, тревогой и ужасом. Между двумя системами существуют реципрокные отношения, так что возбуждение одной из них подавляет деятельность противоположной. Соотношение систем непосредственно регулируется миндалевидным ядром и другими выше расположенными отделами головного мозга.
Гипотеза Боварда подкупает своей стройностью. Давно было отмечено, как велика роль симпатической нервной системы в физиологии отрицательных эмоций. Характерно, что в структуре ориентировочного рефлекса ("биологической осторожности", но И. П. Павлову) только урежение сердцебиений представляет парасимпатическую реакцию. Все остальные проявления ориентировочного рефлекса связаны с активностью симпатического отдела (О. С. Виноградова, 1961). С другой стороны, именно парасимпатической нервной системе принадлежит ведущая роль в деятельности по удовлетворению основных жизненных потребностей организма - пищеварения и функциях полового аппарата. В то время как раздражение заднего ("симпатического") гипоталамуса вызывает у обезьяны реакцию тревоги в сочетании с повышением кровяного давления и учащением сердечного ритма, стимуляция переднего ("парасимпатического") отдела сопровождается успокоением животного, понижением кровяного давления и урежением сердцебиений (Шир и Крогер, 1961). Введение адреналина непосредственно в ретикулярную формацию мозга кошки пробуждает животное и десинхронизирует электроэнцефалограмму. Через 1-2 минуты после инъекции ацетилхолина наблюдаются дремота и синхронизация биоэлектрических потенциалов (Кордеау и др., 1963). Такое типично адренолитическое вещество, как аминазин, подавляя оборонительные реакции животного, не влияет на пищевые (П. К. Анохин, 1957). Интересно, что в психиатрической клинике аминазин дает успокаивающий эффект только в тех случаях, когда основу возбуждения больного составляют отрицательные эмоции агрессии и тревоги. Если возбуждение больного носит положительный характер мании величия, необоснованной радости и т. п., аминазин оказывается неэффективен (Т. Я. Хвиливицкий, 1960).
Однако заманчивая перспектива связать положительные эмоции исключительно с парасимпатической системой, а отрицательные - с симпатической вряд ли соответствует действительности. Многочисленные исследования показывают, что между симпатическим и парасимпатическим отделами существуют гораздо более сложные отношения, чем прямая реципрокность. Нередко нарастание возбуждения симпатического отдела сопровождается одновременным нарастанием активности парасимпатических центров (Гельгорн, 1948, 1959, 1960; П. В. Симонов, 1964а, и др.). В структуре положительных эмоций мы обнаруживаем явно симпатические эффекты, а в структуре отрицательных - парасимпатические. Существует ли какая-нибудь закономерность в этих симпато-парасимпатических отношениях или при возникновении разных эмоций симпатические и парасимпатические реакции комбинируются в самой причудливой последовательности? По-видимому, определенная система все-таки существует.
Небольшой сдвиг вегетативного баланса в парасимпатическом направлении наблюдается при состояниях покоя, комфорта, расслабления. Симпатический сдвиг характерен для бодрости и оптимизма, однако дальнейшее нарастание симпатического тонуса, как правило, уже свидетельствует о напряженности, агрессии и тревоге. Значительное возбуждение парасимпатического отдела мы обнаруживаем при сильном страхе, депрессии и меланхолии (Гельгорп. 1901).
Обращает на себя внимание тот факт, что активно-оборонительные реакции (агрессия, ярость) расположены в "симпатической сфере влияния", а пассивно-оборонительные (страх, депрессия) - в "парасимпатической сфере". По данным И. Т. Джелиева, Н. И. Лагутиной и А. А. Фуфачевой (1963), активно-оборонительные реакции у обезьян сопровождаются тахикардией, а пассивнооборонительные - урежением сердечного ритма. В этом плане становится понятной и парадоксальная для "позитивно-негативной" концепции локализация центров страха в переднем отделе гипоталамуса наряду с реакцией агрессии при раздражении заднего отдела (С. А. Саркисов, 1964).
Рассматривая симпато-парасимпатические отношения при различных эмоциональных состояниях, мы обнаруживаем явное сходство с теми отношениями, которые складываются в вегетативной сфере по мере нарастания силы действующего раздражителя (П. В. Симопов, 1962). Вместе с тем в "ряду эмоций", о котором мы только что говорили, ость одно непонятное звено: преобладание парасимпатического отдела там наблюдается дважды в самом начале "ряда" (покой, комфорт, расслабление) и в самом конце (депрессия, сильный страх). Изучение закономерных изменений вегетативных реакций организма на возрастающий стимул убедило в том, что состояние первичного угнетения существенно отличается от вторичного, запредельного по своей природе. Первичное торможение возникает задолго до того, как нервные структуры исчерпают свои функциональные возможности. При действии достаточно сильного стимула первичное торможение (мы назвали его превентивным) исчезает и сменяется возбуждением. Условия возникновения и биологическая роль превентивного торможения представляют частный случай принципа гиперкомпенсации, о котором мы подробно говорили в предыдущей главе. Весьма вероятно, что преобладание парасимпатической системы при комфорте и удовлетворении реализуется на фоне первичного торможения, а парасимпатические эффекты депрессии и страха выступают па фоне запредельного. Пока трудно сказать, чем отличаются нейрофизиологические механизмы превентивного торможения от механизмов запредельного, с какими структурами связаны эти два вида центрального угнетения. По-видимому, в осуществлении первого из них решающая роль принадлежит кортикальным тормозным аппаратам и неспецифическим ядрам таламуса (Жуве, 1950; А. И. Ройтбак, 1959, 1962). Запредельное торможение связано с более глубокими физико-химическими сдвигами в нервных клетках, в частности с повышением сорбционных свойств клеточной протоплазмы (С. Н. Романов, 1956; Д. Н. Насонов, 1959). Возможно, что при запредельном торможении корковых клеток усиливается тормозящее влияние ретикулярной формации нижнего отдела мозгового ствола. По мнению Моруцци (1902), ото тормозящее влияние сдерживается корой больших полушарий. Мы ужо говорили, что возбуждение симпатического отдела вегетативной нервной системы в большинстве случаев сопровождается активирующим действием ретикулярной формации.
На рис. 3 схематически изображено участие симпатического и парасимпатического отделов вегетативной нервной системы, а также механизмов возбуждения, превентивного и запредельного торможения в осуществлении различных эмоциональных состояний. Схема показывает, что отношения между симпатической и парасимпатической активностью по сводятся к прямой реципрокности. Возбуждение симпатического отдела всегда сопровождается возбуждением парасимпатического отдела, однако "удельный вес" каждой функциональной составляющей специфичен для той или иной группы эмоций. Нельзя не обратить внимания на сходство между схемами рис. 3 и фазными изменениями вегетативных реакций организма по мере нарастания возмущающего воздействия (П. В. Симонов, 1964). Впрочем так ли уж неожиданно это сходство? Рассматривая группы эмоций на рис. 3, мы видим, что для эмоций, расположенных в правой частп схемы, характерны большая степень потребности и больший дефицит информации, чем для состояний, перечисленных слева (согласно формуле, величина эмоции прямо пропорциональна и потребности, и дефициту). Специального анализа требует только место положительных эмоций (второй столбик слева). Их размещение на схеме хороню согласуется с результатами экспериментов.
Мы не раз отмечали, что положительные эмоции по своим вегетативным проявлениям уступают сдвигам в организме, связанным с отрицательными эмоциями, хотя и требуют большей степени напряжения, чем состояние покоя. Но ведь положительные эмоции возникают при избытке информации и но этому признаку должны располагаться слева от покоя. Возникшее противоречие устраняется "формулой эмоций". Наши схемы отражают величину эмоционального напряжения, а не ее знак. Величина Э может достигать значительных размеров и при Н больше С, и при С больше Н. Впрочем при положительных эмоциях возбуждение практически не бывает чрезмерным: науке но известны нервные расстройства от слишком большой радости. Вся клиника неврозов базируется на отрицательных эмоциях, и никто еще не описал психосоматических (кортиковисцеральных) заболевании на почве счастья, удовлетворения и оптимизма. Только в психиатрии мы встречаем патологическое возбуждение позитивных структур (мания величия, эйфория), которое, по-видимому, возникает вторично в результате нарушения высших контролирующих аппаратов.
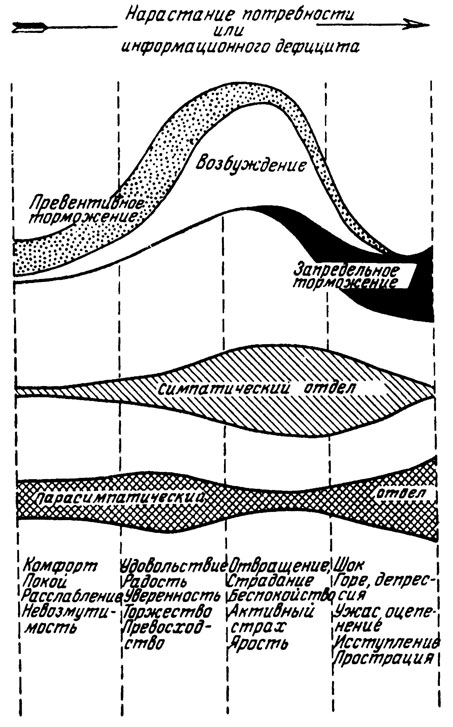
Рис. 3. Схема участия механизмов превентивного торможения, возбуждения и запредельного торможения, а также симпатического и парасимпатического отделов вегетативной нервной системы в реализации различных эмоциональных состояний
Итак, мы рассмотрели пути и закономерности реализации эмоциональных состояний. Теперь необходимо перейти к вопросу о механизмах, которые запускают ту или иную эмоциональную реакцию. Что касается потребностей, то в соответствующей главе были приведены данные о центрах голода, насыщения, жажды, полового влечения и т. д. Но где определяется дефицит информации? Откуда поступают команды к исполнительным центрам разнообразных эмоций?
По мнению И. С. Беритова, регуляция эмоций сосредоточена в древней коре. Именно палоокортекс производит интеграцию активности сомато-вегетативных центров гипоталамуса, центров внешнего выражения эмоций в полноценную эмоциональную реакцию высших позвоночных животных. Среди образований древней коры исследованиями последних лет особое место отводится гиппокампу. Было отмочено, что при решении животным какой-либо задачи (например, поиск пищи в лабиринте) электрическая актин ность гиппокампа усиливается. Она нарастает при ошибках, по исчезает в момент достижения цели (Эйди, 1900-1961; Холмс и Эйди, 1960). Электрическое раздражение гиппокампа ведет к нарастанию ошибок, которые перестают коррегироваться (Прибрам, 1960). Эти экспериментальные факты дали основание рассматривать гиппокамп в качестве аппарата сличения ожидаемого эффекта с реально достигнутым, который П. К. Анохин называет "акцептором действия" (А. Р. Лурия, 1963). Вместе с тем трудно допустить, чтобы сличение происходило в самом гиппокампе. Все данные современной физиологии свидетельствуют о том, что органом накопления индивидуального опыта, нервным устройством замыкания и хранения условных связей является новая кора больших полушарий. Как же тогда понять своеобразие электрической активности гиппокампа? Прежде всего напомним, что гиппокамп относится к числу образований, связанных с отрицательными эмоциями ярости и страха (И. С. Беритов, 1961; Икеда Терутика, 1961; Дельгадо, 1962; Монье-Дюман, 1963 и др.). Неудачи животного при решении задачи и возникающий при этом дефицит информации ведут к развитию отрицательного состояния, которое реализуется с участием гиппокампа. Таким образом, вовлечение гиппокампа в реакцию можно представить себе как явление вторичного порядка, отнюдь не обусловленное процессами сличения в самих гиппокампальных структурах.
Решающая роль в запуске эмоциональных реакции, по-видимому, принадлежит коре больших полушарий. Систематическими исследованиями школы К. М. Быкова, Э. А. Асратяна с сотрудниками и других было показано значение корковых структур для рефляции вегетативной сферы организма. Но данным Фабиана (1961), вегетативный кожногальванический рефлекс возникает у кошек при раздражении лимбической коры. Этот рефлекс зависит от ретикулодермальных влияний, контролируемых корой (Блоч и Бонвалле, 1959). При раздражении новой коры электрические вызванные ответы регистрируются в перегородке, миндалевидном ядро и гиппокампе (Наймер, Гудфеллоу, Спикер, 1963). Особенно тесно связаны с вегетатикой лобные отделы. Лобная кора человека участвует в регуляции сердечно-сосудистых реакций (П. Овчарова, 1962), ее удаление у крыс затрудняет угашение сердечного рефлекса на охлаждение хвоста (Глезер и Гиффип, 1962). Многие факты (мы уже говорили о десинхронизации электроэнцефалограммы, вызванной голодом) дают основание считать, что именно в медно-базальных отделах коры отсутствие положительных факторов (нища), как и действие вредных (боль), создает то состояние напряжения, которое снимается утолением голода или устранением боли. Удаление коры головного мозга существенно изменяет течение эмоции. Реакции декортицированных животных характеризуются гипервозбудимостью и низким порогом вспышек ярости. Двустороннее удаление височной области у обезьян, напротив, приводит к эмоциональной апатии (Линдсли, 1960). Значение корковых структур, хранящих запечатления индивидуального опыта, наглядно демонстрируют опыты Граштьяна, Лишака и Кекеши (1956) с условнорефлекторным переключением. Если у животного один и тот же сигнал в разных условиях подкреплялся пищей или тоном, то раздражение ретикулярной формации вызывает реакцию, соответствующую обстановке. Вне экспериментальной ситуации это раздражение дает только ориентировочный рефлекс. Придание раздражителю сигнального значения делает стойкими вегетативные проявления ориентировочного рефлекса, чего не наблюдается у больных с поражением лобных долей (А. Р. Лурия, 1962).
Клинические наблюдения А. Р. Лурия (1962) и в меньшей мере эксперименты на животных Прибрама (1901 - 19(33) позволяют допустить существование двух кортикальных систем, играющих особо важную роль в организации целенаправленного поведения. По мнению авторов этой концепции, задняя система, расположенная на границе теменной, височной и затылочной областей, путем сопоставления падающих раздражителей со следами впечатлений, полученных ранее, формирует программу действий животного и человека. Передняя префронтальная система контролирует соответствие этой программы с реальным ходом ее выполнения. Действительно, лобная кора расположена очень удачно для осуществления подобных функций. С одной стороны, она тесно связана с "висцеральным мозгом", с центрами органических потребностей живого существа, с нервным аппаратом эмоций. С другой стороны, она непосредственно примыкает к тем областям коры, которые регулируют двигательную сферу, а у человека - и речевые функции. Можно думать, что именно в базальных отделах лобной коры происходит интеграция сигналов из внутренней среды организма с двигательной активностью животного, благодаря чему лобная кора направляет деятельность по удовлетворению жизненных потребностей. Клинические и экспериментальные факты, на которых базируется концепция "внутренних систем коры", делают вероятным предположение об особой причастности лобных и томенно-височных областей к выявлению дефицита информации, необходимой для организации целенаправленного поведения. По-видимому, именно в этих областях на стыке восходящей сигнализации о возникновении потребности и условнорефлекторного опыта, хранящегося в кортикальных структурах, рождаются те нисходящие команды, которые запускают деятельность эмоциональных центров "висцерального мозга". Но для физиолога недостаточно постулировать существование "акцептора действия", "программы", "аппарата сличения" и тому подобных абстракций. Цель физиологии состоит в отыскании нервных механизмов, осуществляющих функции сличения, программирования, динамической коррекции. В этом, решающем для физиологии аспекте мы не располагаем пока никаким реальным багажом, кроме павловского учения об условных рефлексах, в частности о таких формах условнорефлекторной деятельности, как ситуационные условные рефлексы, переключение и динамический стереотип.
Преимущество теории динамического стереотипа состоит в том, что она показывает, как, за счет каких физиологических механизмов формируется "программа" деятельности высших животных. Сформированный динамический стереотип не нуждается в воспроизведении всей системы условных сигналов, соответствующих каждому отдельному звену приспособительного поведения. Опытами Э. А. Асратяна (1953) было убедительно показано, что сложная последовательность реакций может быть осуществлена под влиянием одного и того же раздражителя, который приобретает значение пускового сигнала. Результаты этих опытов демонстрируют еще одно принципиально важное обстоятельство: для сохранения стереотипа решающим фактором оказывается не семантика сигналов (модальность, сила и т. д.), а их прагматическое значение, непосредственно связанное с ответной деятельностью организма. Стереотип условных реакций хорошо воспроизводится раздражителем, который не участвовал в его формировании и очень мало похож на сигналы, образующие систему. Когда вместо шипения, стука метронома, света и звонка мы начинаем применять раздражение кожи касалкой, можно убедиться, что мозг запомнил и сохранил не столько признаки ранее употреблявшихся сигналов, сколько свои реакции на эти сигналы, большие по величине или меньшие, положительные или тормозные. Внутренний стереотип нарушается не при всяком изменении системы внешних раздражителей, а только при таком изменении, которое требует иных, чем прежде, реакций. Именно тогда, т. е. при появлении дефицита информации, необходимой для новых действий, появляются признаки эмоционального напряжения, признаки ориентировочной реакции, представляющей одну из самых распространенных эмоций.
Л. Б. Козаровицкий (1964), исследуя кожногальванический рефлекс у диспетчеров аэропорта, показал, что этот рефлекс угасает, как только заканчивается формирование стереотипа ответных действий, хотя частные особенности падающих раздражителей продолжают изменяться в весьма значительных пределах. В исследованиях П. В. Симонова и М. Н. Валуевой (1963), П. В. Симонова, М. Н. Валуевой и П. М. Ершова (1964) также экспериментально установлена зависимость ориентировочного кожногальванического рефлекса от прагматической, а не семантической функции условных сигналов. Это привело к выводу о том, что ориентировочный рефлекс угасает не в связи с формированием "нервной модели стимула" в мозгу (Е. Н. Соколов, 1960, 1963), а в связи с формированием стереотипа ("модели", "плана", "программы") ответных действий человека или животного. К заключению об активном характере ориентировочной реакции на основании своих экспериментов пришли Н. И. Гращенков, А. М. Вейн, Л. П. Латаш (1963; см. также Н. И. Гращенков и Л. П. Латаш, 1964).
Взаимодействие программирующих (тонических, по Э. А. Асратяну) и пусковых (фазических) сигналов отчетливо выступает в явлениях условнорефлекторного переключения (Э. А. Асратян, 1953,1959). В результате ранее приобретенного опыта раздражитель-переключатель (время опыта, обстановка, вид камеры и т. д.) формирует в коре больших полушарий определенную систему нервных связей повышенной или, наоборот, пониженной возбудимости. Теперь пусковой (фазический) сигнал только включает уже "заготовленную" реакцию, предопределенную условнорефлекторной "программой". Переключение занимает как бы промежуточное положение между динамическим стереотипом с постоянными интервалами между раздражениями, образующими систему, и ситуационным условным рефлексом (доминантой). В случае стереотипа заранее определен не только характер, но и момент ответной реакции. При переключении живая система заблаговременно информируется о необходимых действиях (овладение нищей, избегание тока и т. д.), но не имеет сведений о том, когда их надо совершить. Начало действия определяется пусковым раздражителем.
Таким образом, в ситуацию переключения вводится некоторая доза неопределенности, что и влечет за собой элементы эмоционального напряжения, связанного с готовностью ответить на пусковой раздражитель, когда бы он ни появился. Однако переключение - еще не доминанта, потому что влияние тонического сигнала избирательно распространяется лишь на заранее выработанные условнорефлекторные связи.
Но вернемся к ситуации нарушенного стереотипа, когда изменения в окружающей среде сделали неэффективной существующую последовательность действий. Здесь-то и включаются механизмы эмоционального реагирования, призванные скомпенсировать "информационные пустоты" до тех пор, пока не сформируется новый стереотип. Факт вовлечения нервного аппарата эмоций мы обнаруживаем по многочисленным вегетативным сдвигам в организме, по изменению электрической активности подкорковых образований. Ранняя стадия выработки условного рефлекса (или новой системы условнорефлекторных связей) требует участия лимбической или гиппокамиальной системы, тогда как позднее преобладает активность структур новой коры (Моррелл, Барлоу, Брейзье, 1960). Это не следует понимать в том смысле, что новый условный рефлекс начинает формироваться в подкорке, а потом "переходит" в кору. Процесс инициируется корой больших полушарий, но на первых этапах кора особенно активно привлекает к участию подкорковые образования с присущими им формами широко генерализованного доминантного реагирования. Ярким примером кортиколизованных доминант могут служить ситуационные условные рефлексы, когда отдаленный условный сигнал создает пищевую, половую или оборонительную доминанту. Теперь любой, даже впервые встретившийся в жизни раздражитель будет вызывать реакцию, соответствующую доминанте. Так, разведчик, войдя в незнакомый лес, хватается за автомат при каждом шорохе, движении, мелькнувшей тени. Нетрудно понять, насколько выгодно такое гиперкомпенсаторное поведение в обстановке, когда потребность в защите сочетается с острой нехваткой сведений о путях ее надежного обеспечения.
Главный результат вовлечения нервного аппарата эмоций состоит в переходе живой системы к доминантным типам реагирования, благодаря которому индифферентные раздражители или сигналы, ранее имевшие другое значение, профилактически начинают вызывать господствующую (ориентировочную, оборонительную, половую) реакцию. Возникшая доминанта подавляет и отменяет другие, в данный момент второстепенные деятельности организма, сосредоточивает все силы и резервы на удовлетворении наиболее острой потребности. Эмоциональные доминанты обладают выраженной инерционностью, они обеспечивают продолжение деятельности даже при хронической неэффективности двигательных реакций, в обстановке, когда достижение цели кажется маловероятным. Тем самым эмоциональное возбуждение способствует поиску недостающей информации, представляет движущую пружину этого поиска. Мы уже говорили о мощном активирующем влиянии эмоций на высшие отделы мозга, которое в значительной мере реализуется через ретикулярную формацию мозгового ствола. Эмоция регулирует уровень бодрствования, уровень работоспособности коры больших полушарий. Восходящие тонизирующие влияния способствуют мобилизации опыта, хранящегося в условно- рефлекторных связях коры (Гис, 1963). Частный случаи подобной мобилизации высшего порядка мы наблюдаем у человека в состоянии вдохновения, когда возникают столь неожиданные догадки и ассоциации.
Через симпатический и парасимпатический отделы вегетативной нервной системы усиливается деятельность органов сердечно-сосудистой системы, дыхания, внутренней секреции. Это усиление вполне понятно, если учесть, что доминантное гиперкомпенсаторное поведение всегда расточительно. Оно не позволяет обеспечить строгое соответствие двигательных и вегетативных реакций, как это наблюдается в случаях динамической стереотипии. Вегетативные сдвиги при эмоциях представляют энергетическое обеспечение с запасом, с поправкой на непредвиденные обстоятельства.
Все перечисленные нами характеристики эмоционального реагирования продуктивны и целесообразны только в условиях информационного дефицита. По мере ликвидации этого дефицита, по мере формирования нового динамического стереотипа достоинства эмоций диалектически превращаются в недостатки. Реакции, точно и тонко соответствующие внешним сигналам, гораздо эффективнее доминантного поведения. Сильное возбуждение кортикальных структур способно только помешать осуществлению двигательного навыка, а расточительность вегетативных сдвигов существенно нарушает энергетический баланс организма. Теперь мы видим другую сторону эмоций: когда излишнее волнение мешает спортсмену показать на ответственных соревнованиях те результаты, которых он добивался на тренировках; когда человек совершает нелепые и опрометчивые поступки; когда возникают неврозы - патологические расстройства психоэмоциональной сферы.
До сих пор мы говорили о том общем, что присуще самым разнообразным эмоциям высших животных и человека. Настало время перейти к физиологической классификации эмоциональных состояний.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://psychologylib.ru/ 'Библиотека по психологии'