§ 2. Активирующая ретикулярная формация
В основании мозга, в центральной области ствола, от продолговатого мозга до гипоталамуса простирается структура, издавна известная анатомам под названием ретикулярной формации (рис. 2). Она получила это название из-за своего сетчатого строения, поскольку она состоит из значительного числа нервных клеток (в ней содержится 9/10 клеток зсего ствола мозга), сплетенных и связанных между собой сетью соединительных клеток (рис. 3). Ретикулярная формация, которая существует у всех позвоночных и значение которой возрастает соответственно месту животного в эволюционном ряду, до недавнего времени была terra incognita на карте нервной системы. Современные исследования показали, что эта структура представляет собой нечто вроде усилителя и распределителя нервной информации. Эта информация поступает из внешней среды в форме центростремительных нервных импульсов от органов чувств и из внутренней среды через посредство проприо-цепторов и интероцепторов; она создается также стимуляцией, вызываемой изменениями химического состава крови - увеличением количества адреналина (Бонвалле, Делл и Хибел, 1954) или углекислого газа (Южлен, Бонвалле и Делл, 1959), циркулирующего в организме. Затем она возвращается в организм в форме центробежных нервных импульсов, которые оказывают облегчающее влияние на двигательную, вегетативную системы и кору мозга (Понятия сенсорной и ретикулярной динамогении не предполагают, что нервная система получает энергию в собственном смысле слова из внешней среды и передает ее эффекторным органам. Нервная система передает не энергию, а информацию в форме цифрового кода (частота импульсов), и эта информация вызывает в пункте ее назначения облегчающее или тормозное влияние. Когда говорят, что активирующая ретикулярная формация является неспецифической системой, этим хотят сказать только, что эта структура, по-видимому, особенно чувствительна к общему количеству импульсов, поступающих к ней, и менее чувствительна к формальному содержанию сообщений. Однако, как мы увидим в дальнейшем, значение некоторых особенно важных сообщений может быть распознано уже на уровне подобных структур).
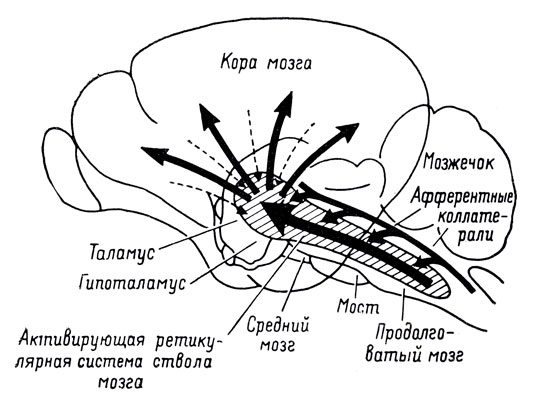
Рис. 2. Схематическое изображение мозга кошки, показывающее расположение ретикулярной формации, распределение коллатералей афферентных путей и активирующие кортикальные проекции (по Старцлу Т. Э., Тэйлору С. У. и Мэгуну Г. У.)
Действительно, можно показать, что:
а) Активирующая ретикулярная формация (АРФ) является местом конвергенции всех сенсорных путей. От каждого рецептора, например глаза, уха, тактильных клеток, рецепторов натяжения и движения мышц и т. д., отходят нервные волокна, которые после определенного числа реле-ядер достигают ре-цепторных зон коры: по этим волокнам передаются нервные импульсы, которые информируют нас о любом изменении внешней среды или нашего положения по отношению к ней. Те же самые волокна, когда они проходят на уровне ствола мозга, дают разветвления, уходящие в массу ретикулярной формации и называемые коллатералями (рис. 3). Электрофизиологическая техника позволила показать, что в одном и том же пункте этой структуры и даже на одном и том же нейроне могут конвергировать импульсы, поступающие от рецепторов разной модальности. Эги импульсы, идущие по коллатералям, по-видимому, теряют свой специфический характер и выполняют информационную роль в АРФ только посредством своего общего объема. Мы можем, таким образом, представить себе, что всякая стимуляция, воздействующая на организм, имеет двоякое назначение: она доставляет специфическую информацию по специфическим путям и одновременно увеличивает общий количественный показатель через посредство такой рассеянной структуры, какой является ретикулярная формация.
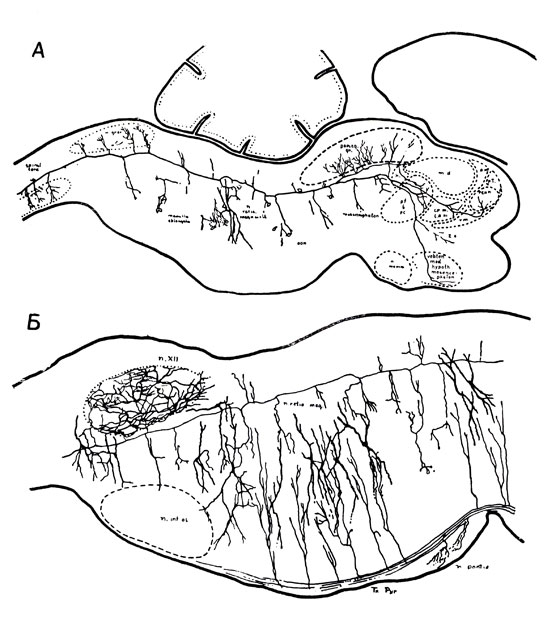
Рис. 3. А. Нейрон ретикулярной формации. Сагиттальный разрез ствола мозга крысенка. На рисунке представлен только один нейрон ретикулярной формации. Видно, что аксон разделяется на каудальный и ростральный сегменты, большой протяженности и со множеством коллатералей. Б. Коллатерали. Сагиттальный разрез нижней части ствола мозга крысенка, показывающий соединения коллатералей большого нисходящего пути (пирамидный путь) с ретикулярными нейронами. Коллатерали восходящих путей (сенсорные пути), отсутствующие на рисунке, соединяются с ретикулярными нейронами подобным же образом (по Шейбэлу М. Э. и Шейбэлу А. Б.)
б) В зависимости от количества импульсов, поступающих по коллатералям специфических сенсорных путей, и химических изменений внутренней среды ретикулярная формация обнаруживает различные уровни активности, которые многие психологи называют изменениями ретикулярного тонуса.
Эти изменения ретикулярного тонуса обнаруживаются по их влиянию на активность мозговой коры и возбудимость двигательных и вегетативных систем.
в) Действительно, активность ретикулярной формации оказывает генерализованное облегчающее влияние. Аксоны ретикулярных нейронов образуют восходящие эфферентные пути к мозговой коре и нисходящие к мотонейронам и симпатическим эффекторным нейронам мозга (рис. 3).
Восходящие ретикулофугальные пути диффузно проецируются на всю мозговую кору. Следствием такой проекции, которая захватывает все зоны коры, является то, что всякое повышение ретикулярного тонуса влечет за собой общую активацию коры. Эта активация при электроэнцефалографической регистрации выражается в классической "реакции блокады" или реакции пробуждения, то есть в исчезновении медленных волн большой амплитуды, характерных для сна и состояния покоя, и замене их волнами высокой частоты и малой амплитуды.
Нисходящие ретикулофугальные пути осуществляют облегчающее влияние на мотонейроны спинного мозга и вызывают, следовательно, снижение порога двигательных реакций и повышение позного тонуса. Наконец, эти пути проецируются также на спинномозговые нейроны симпатической системы, и ретикулярная активация вызывает ускорение сердечного ритма, повышение артериального давления, расширение зрачка, электрокожные разряды и т. д. Важно подчеркнуть, что все эти облегчающие влияния оказываются одновременно и исходят из одних и тех, же областей ретикулярных структур, составляющих, таким образом, единую функциональную систему (В результате первых экспериментов Мэгуна и Райнеса было высказано предположение о существовании двух активирующих систем: восходящей активирующей системы, оказывающей влияние на кору мозга, и нисходящей активирующей системы, облегчающей функционирование двигательных нейронов. Однако данные экспериментальных работ Южлена и Бонвалле (1957) позволили сделать вывод, что восходящие, облегчающие влияния на кору и нисходящие влияния на двигательную систему осуществляются одними и теми же областями ретикулярной формации. Кроме того, Блоку и Бонвалле (I960) и Бонвалле и Зброзине удалось доказать, что раздражение одних и тех же областей этих формаций оказывает одновременное действие на кору, двигательную и симпатическую системы). Это значит, что ретикулярная активация вызывает одновременно увеличение активности коры мозга, снижение порогов двигательных реакций и усиление симпатических функций. В плане поведения подобный процесс способствует более многообразному и богатому взаимодействию между организмом и внешним миром: иначе говоря, он ведет к повышению уровня бодр* ствования.
г) Существует множество экспериментальных доказательств роли активирующей формации в регуляции бодрствования.
В то время как при экспериментальных перерезках специфических сенсорных путей к коре мозга реакция пробуждения у животных еще возникает, обширные повреждения АРФ погружают их в состояние оцепенения, сопровождающегося ЭЭГ, характерной для непрерывного сна. Установлено также, что снотворные агенты (гарденаль, барбитураты) воздействуют именно на ретикулярную формацию.
С другой стороны, у спящих животных электрическая стимуляция АРФ вызывает пробуждение, сопровождающееся картиной активации на ЭЭГ и облегчением двигательных и вегетативных функций. Важно отметить, что это влияние аналогично тому, которое можно получить посредством достаточно интенсивного раздражения специфических сенсорных путей (см. рис. 4).
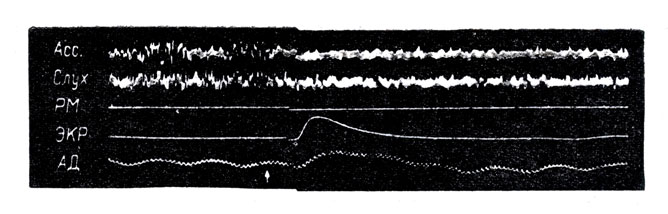
Рис. 4. Одновременное воздействие сенсорной стимуляции на кортикальные двигательные и вегетативные функции. Слуховой стимул (щелчок) вызывает реакцию пробуждения на электрокортикограмме (ассоциативная и слуховая кора), облегчение моносинаптического рефлекса (РМ), электрокожной реакции (ЭКР), повышение артериального давления (АД). Все эти влияния стимула на активирующую ретикулярную формацию осуществляются через посредство коллатералей центральных чувствительных путей. Прямое раздражение ретикулярной формации влечет за собой аналогичный эффект. (По Блоку В.)
Такое ретикулярное возбуждение можно вызывать в "хроническом" опыте у бодрствующих и свободно передвигающихся животных с помощью предварительно вживленных электродов, связанных с аппаратами стимуляции и энцефалографом либо посредством длинных проводов, либо радиоэлектрическим путем. Одновременный анализ поведения и ЭЭГ таких животных представляет значительный интерес для изучения регуляции бодрствования. В самом деле, если очень постепенно увеличивать интенсивность стимуляции начиная с очень слабой, то можно получить у животных всю последовательность поведенческих реакций, от сна до ярости в соответствии с континуумом, о котором шла речь выше. Последовательно наблюдается приподнимание головы, открывание глаз, исследовательские движения головой, выпрямление тела, поисковые перемещения; затем эта активность усиливается и, когда уровень возбуждения достигает высоких значений, превращается в беспорядочное возбуждение, сопровождающееся признаками вегетативного расстройства.
Эти наблюдения показывают, что одни и те же области нервной системы - в зависимости от степени их возбуждения - могут определять изменения в уровне бодрствования, обусловливающие столь различное поведение, как простое пробуждение, реакция внимания без определенной цели, эмоциональная разрядка. Эти нервные структуры, по-видимому, ответственны за изменения поведения по интенсивности, сопровождающие изменения, связанные с его направленностью (см. стр. 97).
Поэтому при изложении мы будем исходить из того, что существует единая шкала бодрствования, на которой пробуждение от сна и мобилизация внимания (и мы могли бы добавить эмоциональное пробуждение) представляют собой переход от одного уровня бодрствования к другому в соответствии со степенью ретикулярной активации. В согласии с этой схемой мы последовательно остановимся на таких проблемах, как сон, пробуждение и уровни бодрствования, пробуждение внимания и внимание как процесс.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://psychologylib.ru/ 'Библиотека по психологии'