§ 3. Концентрация внимания
Проблема внимания не ограничивается только пробуждением внимания, которое, как мы видели, вызывает генерализованное облегчение и является лишь подготовительным для специфической деятельности. Все психологи подчеркивают то свойство внимания, которое состоит в концентрации его на одной воспринимающей или эффекторной сфере и предполагает торможение конкурирующих сфер.
Первый этап такой концентрации может осуществляться уже на уровне органов чувств, по крайней мере в отношении зрения, которое у человека имеет первостепенное значение. В самом деле, известно, что рефлекс фиксации имеет своим назначением помещение на фовеальную область (центральную область сетчатки, где разрешающая способность максимальна) образа предмета, изменения которого, воспринимаемые периферическим зрением, и составляют пробуждающие внимание стимулы.
Мало что известно о механизмах этого важного рефлекса, кроме того, что он вводит в действие высшие мозговые структуры и что его онтогенетическое формирование является сравнительно продолжительным (Лезевр, 1964). Из-за ограниченности фовеальной части, позволяющей при одной фиксации воспринять лишь небольшое число объектов, фиксация представляет собой исключительный пример перцептивной концентрации. Действительно, направить свое внимание на один зрительный стимул -это значит прежде всего его фиксировать. Вот почему изучение фиксационных точек, порядка, в котором они следуют, и особенно длительности каждой из последовательных фиксаций составляет прямой способ оценки зрительного внимания. Такое изучение стало возможным благодаря регистрации движений глаз, устанавливающей место и продолжительность фиксаций, которые можно перенести на точки объекта-стимула. Такими методами являются либо фотографическая регистрация угловых смещений тонкого светового пучка (видимого или инфракрасного), отраженного от роговицы или зеркала, закрепленного на глазу, либо кинематографическая регистрация движений глаза, либо регистрация изменений потенциалов, связанных с движением глаз (электроокулография) (см. рис. 13 и 14).

Рис. 13. Внизу. Запись движений глаз (зарегистрированных оптическим методом), соответствующих зрительному изучению предъявленного рисунка (наверху). Обратите внимание на частоту фиксаций на интересных точках и обусловленность взгляда структурой объекта - стимула (по Ярбусу)
Однако у опытного испытуемого возможна диссоциация между зрительной фиксацией и вниманием благодаря произвольной диплопии, при которой фиксируемая точка проецируется в центр двух фовеа, тогда как точка, на которую направлено внимание, проецируется на некорреспондирующие точки поверхности сетчатки. Трудность этой операции как раз и свидетельствует о том, насколько функционально связаны между собой фиксация и внимание.
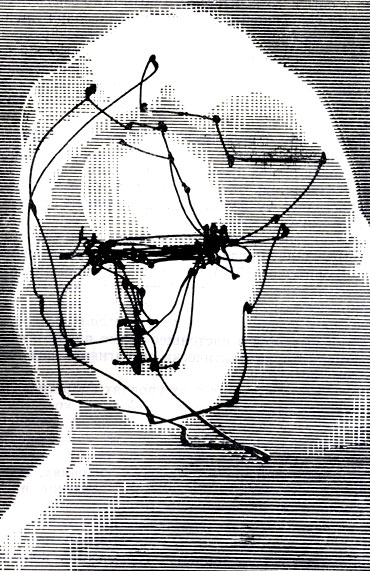
Рис. 14. См. подпись к рис. 13
Этот излюбленный пример из области зрения служит хорошей иллюстрацией двойного аспекта внимания: периферический стимул вызывает общее пробуждение, к которому присоединяется рефлекс фиксации; затем фиксация в собственном смысле слова осуществляет ту функцию концентрации, которая характеризует внимание.
Но даже в этом случае "перцептивная центрация" ("centration perceptive") не всегда совпадает с направлением взгляда, и Фресс и Вюрпилло (1956) показали, что фиксация взгляда и направленность внимания могут быть разъединены в этом явлении. Избирательная направленность внимания предполагает, следовательно, в сущности, процессы центральной природы. Нейрофизиология пытается в настоящее время подойти к этой трудной проблеме, которая фактически затрагивает еще более сложную проблему сознания.
Мы уже отмечали, что на всех уровнях бодрствования можно получать в реле-ядрах сенсорных путей или в коре мозга вызванные потенциалы, которые отражают движение сенсорных сообщений, их приход или преобразование на уровне центров. Возникает вопрос, может ли изучение этих показателей дать представление о том, как при всех этих сообщениях, достигающих центров, осуществляется избирательное усиление привилегированного сообщения, объясняющее концентрацию внимания.
Можно предложить две гипотезы: либо существует особый механизм, усиливающий действие пробуждающего внимание стимула, либо имеются механизмы торможения конкурирующих стимулов.
А. Усиление действия стимула, пробуждающего внимание
Обнаружено, что у животных и человека при постоянных стимулах вызванные потенциалы обнаруживают изменения амплитуды и области распространения, связанные с изменениями внимания, оцениваемыми самостоятельно или вызываемыми экспериментально.
Литература по этому вопросу хотя и обширна, но довольно разноречива, поскольку вызванные потенциалы крайне сложны как феномен и являются результатом взаимодействия многочисленных факторов. Однако, если попытаться выбрать среди различных исследований лишь те, которые действительно сравнимы, можно резюмировать данные, собранные в настоящее время этим методом, следующим образом.
В проекционных областях коры зарегистрированные вызванные потенциалы включают в себя по крайней мере два ряда компонентов: колебания с коротким латентным периодом, соответствующим синаптическому потенциалу на уровне корковых клеток, и более поздние модификации дендритных потенциалов, отражающие приход сенсорного сообщения. Недавно было доказано (Дебекер, Демед и Маниль, 1965), что у человека этот первичный потенциал появляется только тогда, когда вызывающий его стимул воспринимается не менее, чем в 25% предъявлений. Но если первичный вызванный потенциал может быть, таким образом, в оптимальных условиях приведен в соответствие с перцептивными порогами, то другие компоненты коркового ответа обнаруживают колебания, связанные с вниманием. Действительно, за первичными компонентами ответов позднее следует совокупность компонентов сложной формы, гораздо более подверженная флуктуациям; она может быть зарегистрирована на уровне проекционных зон коры (неспецифические или "ретикулярные" ответы Брейзье, 1964), а также ассоциативных корковых областей ("вторичные", Бюзе и Боренштейн, 1957, или "ассоциативные" ответы у животных, Альбе-Фессар и Ружель, 1958). Эти компоненты ответа увеличиваются по амплитуде, когда субъект направляет свое внимание на вызвавший его стимул. Демед и сотр. показали, что если испытуемый получает инструкцию сконцентрировать свое внимание на тактильном стимуле (мысленно считая слабые удары по пальцу), соместетический вызванный потенциал обнаруживает вторичные компоненты большой амплитуды; последние исчезают, когда испытуемого отвлекают одновременным предъявлением слуховых стимулов (см. рис. 15). Повышение внимания, направленного на звуки, когда нужно оценить их различительную интенсивность, также влечет за собой увеличение слухового вызванного потенциала (Дэвис, 1964). Наконец, эти флуктуации вызванных потенциалов, по-видимому, обнаруживают корреляцию с изменениями уровня бодрствования, оцениваемого по выполнению психометрических задач большой продолжительности: отключения внимания, устанавливаемые по необнаружению зрительного сигнала, выражаются в сокращениях амплитуды зрительных вызванных потенциалов (Хайдер, Спонг и Линдсли, 1964).
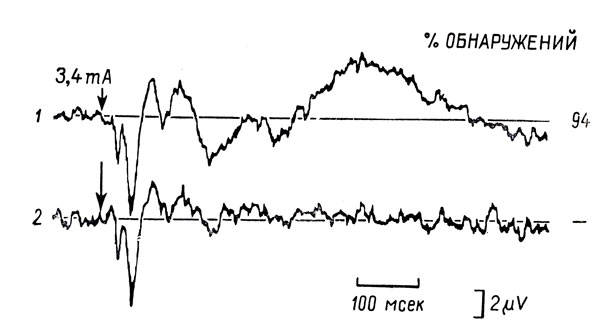
Рис. 15. Модификация соместетического вызванного потенциала в результате концентрации внимания. Представлено усреднение 200 ответов, автоматически произведенное аппаратом с электронной памятью. Стимулом служил короткий электрический удар 3,4 ма, наносимый на палец. 1. Испытуемому дается инструкция сосредоточить внимание на своих ощущениях и мысленно считать удары (процент обнаружений указан на рис. справа). 2. Испытуемому дается инструкция считать слуховые щелчки, предъявляемые по второй независимой программе, в ходе всего сеанса. Отметьте наличие позднего компонента соместетического потенциала большой амплитуды в ситуации 1 (концентрация внимания) и его исчезновение в ситуации 2 (отвлечение). (По Демеду Дж. Э., Дебекеру Дж. и Манилю Дж.)
Вторичные кортикальные вызванные ответы обнаруживают топографические изменения, которые, вероятно, говорят об их распространении на большую территорию коры, когда субъект направляет свое внимание на стимул. Гарсия-Ауст, Богач и Ванцулли (1963) действительно констатировали такое распространение, когда испытуемый должен был сравнивать световые вспышки. Это распространение, в сущности, состоит в том, что ответы не ограничиваются первичными рецепторными зонами, а регистрируются также и в ассоциативных зонах коры. Эти исследования показывают, таким образом, что внимание проявляется в повышении активности затронутых стимулом кортикальных зон, а также в захвате афферентным сообщением обширных территорий коры. Следовательно, "фокусировка" как образ не объясняет центральных явлений. Внимание с перцептивной точки зрения играет роль диафрагмы, в то же время оно вызывает мобилизацию центров в пользу стимулов, привлекающих внимание. Как понять это усиление действия привлекающих внимание сообщений? Объясняется ли оно непосредственно активацией структур, регулирующих уровень бодрствования? Этот последний вопрос в настоящее время очень запутан противоречивостью данных, полученных в результате нейрофизиологических экспериментов. Действительно, если обратиться не к поведенческим критериям внимания, а к изменениям в ЭЭГ, то обычно корковые вызванные потенциалы (первичные или вторичные) больше по амплитуде на фоц? "десицхронизации" в ЭЭГ, то есть при высоком уровне бодрствования. Наряду с этим ретикулярная стимуляция у животных влечет за собой депрессию вызванных потенциалов. Это противоречие нередко ставило в тупик нейрофизиологов. Однако следует напомнить несколько фактов, которые, возможно, прольют некоторый свет на механизмы внимания.
В момент кортикального пробуждения вызванные потенциалы могут обнаруживать большую амплитуду, если вызывающий их стимул ответствен также и за реакцию пробуждения. То, что вызывает внезапное пробуждение внимания, одновременно влечет за собой, следовательно, диффузную кортикальную активацию и значительное усиление сенсорных сообщений. Продолжительность такого облегчения была определена для первичного потенциала Дюмон и Деллем (1960) в 100-250 мсек, период, совпадающий с моторными проявлениями ориентировочной реакции. Дюмон и Делл (1960), а также Бремер, Ступел и Ван-Рет (1960) показали, что если первичные кортикальные потенциалы вызываются не адекватным для них стимулом, а раздражением электрическим током сенсорных путей, то ретикулярная стимуляция вызывает значительное облегчение этих потенциалов. Предполагают, что, если стимул достаточно интенсивен, сенсорное сообщение сможет преодолеть рефрактерность кортикальных нейронов, возбужденных неспецифическими активирующими афферентными раздражителями ретикулярной природы, действие которых выражается на ЭЭГ реакцией пробуждения. Влияние ретикулярной активации на кортикальные нейроны является, по всей видимости, двойственным: с одной стороны, оно снижает порог их ответа на специфические сообщения, усиливает последние и облегчает внутрикортикальную передачу; с другой - оно повышает их автономную активность, которая выражается в реакции пробуждения. Факт конвергенции на одних и тех же нейронах специфических и неспецифических импульсов объясняет то, что если последние слишком значительны, то они вызывают замыкание нейронов, которые становятся рефрактерными к специфическим сообщениям. Этот последний феномен можно продемонстрировать с помощью инъекции амфетамина (Бремер и Ступел, 1959) - наркотика, который повышает ретикулярный тонус, вызывает интенсивное пробуждение и снимает облегчение вызванных потенциалов. Впрочем, часто отмечалось, что употребление этих возбуждающих средств в значительных дозах хотя и повышает общий уровень активности, влечет за собой нарушения бодрствования. Мы сталкиваемся здесь с понятием "оптимума активации", часто упоминаемого в плане поведения.
Бремер и сотр. (1960) предложили вероятностное истолкование этих проблем: ретикулярная активация средней интенсивности, по-видимому, исключает из ответа на специфические импульсы некоторое число корковых нейронов. Но в то же время она усиливает, вероятно, способность передачи самих специфических сенсорных импульсов и увеличивает внутримозговую иррадиацию нейронов, возбужденных этими импульсами. Таким образом, по всей видимости, реализуется этот избирательный, концентрирующий аспект, исчезающий, когда активация слишком интенсивна: в этом случае преобладание тормозного влияния объясняет явления сенсорного "ослепления", характеризующего состояния крайнего эмоционального возбуждения.
Все предыдущие соображения касаются возможного взаимодействия коры и ретикулярной формации в явлениях концентрации внимания. Однако для объяснения этих явлений ссылаются также на возможное вмешательство структуры, которая, по-видимому, ответственна за специфическое бодрствование (Сулерак, 1958) и активность которой накладывается на активность, регулирующую диффузное бодрствование. В самом деле, неспецифическая таламическая система (или таламическая ретикулярная формация) представляет собой ростральное продолжение активирующей ретикулярной формации, но свойства ее другие: стимуляция ее вызывает более локальную реакцию пробуждения в коре (Джаспер, 1949), а ее разрушение уничтожает при выработке условного рефлекса ту фазу, когда у нормального животного условный сигнал вызывает локальное кортикальное пробуждение в зоне условного раздражителя (Моррелл, 1953); наконец, в отличие от активирующей ретикулярной формации (структуры, чувствительной к изменениям количества адреналина в крови) функционирование неспецифических таламических ядер имеет, по-видимому, холинэргическую природу (Ринальди и Химвич, 1955). Эта последняя черта роднит эту структуру с парасимпатической системой, влияния которой, как известно, более специфичны и локальны, чем влияния симпатической системы. Однако явление рекрутмента, или реакция вовлечения кортикальных ритмов, вызываемое низкочастотной стимуляцией таламической ретикулярной формации (Демпси и Морисон, 1942), по-видимому, имеет отношение к дифференциации подкрепляемых и неподкрепляемых стимулов при выработке условных рефлексов. В самом деле, Вайнбергеру и сотр. (1965) удалось модифицировать этот ответ при предъявлении подкрепляемого стимула, тогда как предъявление неподкрепляемого стимула его не изменяло. Можно, следовательно, предположить (Фессар и Гасто, 1958), что стимул, привлекающий внимание, влечет за собой сначала генерализованное пробуждение ретикулярной формации, а затем - на втором этапе - приводит в действие таламическую ретикулярную формацию, которая связана уже с более специфическими влияниями. Понятно, однако, что в настоящее время подобные схемы остаются весьма гипотетическими.
Б. Торможение конкурирующих стимулов
Вместо того чтобы искать механизмы, ответственные за усиление сенсорных сообщений, передающих информацию, содержащуюся в стимуле, привлекающем внимание, можно предположить, что концентрация внимания является результатом торможения сообщений, исходящих от других, стимулов. Ряд экспериментов, проведенных Жуве (1960) и Эрнандес-Пеоном (1961), привел этих авторов к предположению о существовании специального аппарата контроля, пропускающего к коре лишь значимые сообщения, в то время как другие блокируются на периферии. У кошки, например, слуховые потенциалы, зарегистрированные на уровне периферического реле-ядра (кохлеарное ядро), могут исчезнуть, если внимание животного направлено на такой стимул, как мышь (Эрнандес-Пеон, Шеррер и Жуве, 1956). Жуве (1957) получил у человека уменьшение амплитуды зрительных вызванных потенциалов в наружном коленчатом теле, когда субъект направлял свое внимание на незрительные стимулы.
Подобный контроль центров над периферией не представляет собой ничего неправдоподобного, и многочисленные анатомо-физиологические данные говорят о существовании центрифу-гальных путей, идущих либо к сенсорным реле-ядрам, либо к самим рецепторам. Между тем истолкование упомянутых и им подобных экспериментов неоднократно подвергалось сомнению (см., например, Южлен, Дюмон и Пайа, 1960; Хори, 1965), поэтому эту проблему не следует считать окончательно решенной. Во всяком случае, не известно, какой механизм лежит в основе такого избирательного тормозного контроля.
В конечном счете концентрацию внимания можно по-видимому, понимать как результат процесса контраста, основанного на природе стимулов, привлекающих внимание. Если допустить, что эти стимулы обладают качествами интенсивности, новизны и аффективного "заряда", которые в данный момент отличают их от других стимулов, то можно считать, что пробуждение внимания к ним закономерно. Дж.-А. Дейч и Д. Дейч (1953) отметили в связи с этим, что необязательно ссылаться на аналитическую систему, которая сравнивает якобы относительное значение всех сообщений по отношению друг к другу: действие всякого стимула зависит только от непосредственно предшествующего ему состояния систем регуляции бодрствования. Наконец, важные работы Экклса (1965) о пресинаптическом торможении позволяют сделать вывод об отборе сообщений даже до того, как они преодолеют синапсы вставочных нейронов на уровне коры мозга или ретикулярной формации. "Важное" сообщение, то есть сообщение, имеющее большую интенсивность по сравнению с другими стимулами, действующими в данный момент, способно устранить влияние этих последних на центральную нервную систему. В этом случае избирательность и концентрация внимания осуществляются самой рецепторной системой без вмешательства центрифугального тормозного механизма.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://psychologylib.ru/ 'Библиотека по психологии'