V. Состояние бодрствования
§ 1. Пробуждение
Согласно ранее предложенной схеме, переход от одного уровня бодрствования к другому, более высокому, становится возможным благодаря активации нервных процессов. В случае перехода от состояния сна к состоянию бодрствования эта активация проявляется в том, что в обыденном языке называется пробуждением.
Это пробуждение может происходить по видимости спонтанно: в настоящее время нам почти совсем не известны внутренние механизмы, которые могут вызывать пробуждение, особенно когда оно обусловлено временем. Но когда пробуждение явно вызывается внешними стимулами (или даже внутренними, например сокращениями желудка), мы можем попытаться проанализировать природу пробуждающих стимулов. Так, во время сна организм продолжает подвергаться непрерывному потоку раздражений. С помощью метода вызванных потенциалов (см. дальше) можно проследить движение многих сенсорных сигналов вплоть до самой коры мозга; однако они не вызывают изменений в уровне сна. Следовательно, из всех стимулов, продолжающих оказывать свое действие на высшие центры, только некоторые способны разбудить организм. Мы можем задать вопрос, каковы те условия, которые необходимы, чтобы вызвать пробуждение. Эти условия можно, по-видимому, свести к трем главным признакам: интенсивность, новизна и мотивационный или аффективный "заряд" (значение), то есть те самые признаки, с которыми мы встретимся при описании стимулов, способных вызвать пробуждение внимания.
Качества пробуждающих стимулов:
А. Интенсивность
Определенный уровень интенсивности пробуждающего стимула на первый взгляд кажется самым важным условием его эффективности. К тому же именно на этом качестве основан один из критериев оценки глубины сна: сон считается тем глубже, чем выше порог интенсивности, необходимый, чтобы вызвать пробуждение (Клейтман, 1963).
В плане физиологических механизмов правомерно предположить, что для повышения ретикулярного тонуса, сильно угнетенного во время сна и подверженного тормозным влияниям, необходима значительная интенсивность стимуляции. Понятно также, что может быть достаточно одного этого признака интенсивности независимо от значения стимула, чтобы вызвать ретикулярную активацию через посредство коллатералей сенсорных путей.
Однако опыт показывает, что этот признак, каким бы важным он ни был, не составляет основного условия пробуждающей стимуляции, и повседневные наблюдения изобилуют примерами пробуждений, вызванных очень слабыми стимулами, так же как и сна, не нарушаемого очень сильными раздражителями. Поэтому, по-видимому, важнее, два других качества стимула.
Б. Новизна
Сюда мы относим случаи, когда в сенсорном окружении спящего субъекта происходят изменения. В самом деле, если признаки стимулов мало изменяются или изменение их интенсивности происходит постепенно, сон продолжается. Напротив - и это широко известный факт, - всякое внезапное изменение всей стимуляции повлечет за собой пробуждение.
В этом случае физиология ретикулярной формации также может снабдить нас данными, позволяющими объяснить этот феномен. Ретикулярная формация в состоянии бодрствования подчинена тормозному контролю со стороны коры больших полушарий, составляющему часть кортико-ретикулярной цепи обратной связи (Южлен и Бонвалле, 1957). Именно ретикулярный разряд, вызывая генерализованную активацию коры, запускает и регулирует корковые механизмы, которые в свою очередь угнетают ретикулярный тонус. Во время сна ретикулярный тонус низкий (По крайней мере при "классическом" сне (см. стр. 112)) и, следовательно, кортикальный тонус также. Когда интенсивность стимула увеличивается постепенно, ретикулярный тонус повышается медленно, и параллельное медленное повышение кортикального тонуса влечет за собой постепенное распространение торможения и противодействует, таким образом, ретикулярной активации моторных и вегетативных функций (Блок и Бонвалле, 1959). Но если стимул оказывает свое действие внезапно, ретикулярная активация протекает слишком быстро, чтобы быть контролируемой кортикальным пробуждением, и, ничем не сдерживаемая, вызывает полное пробуждение коры и облегчение двигательных и вегетативных функций (см. рис. 7). Характер ответа будет, следовательно, зависеть от временной константы кортико-ретикулярной системы, причем система является стабильной, если входной сигнал обнаруживает медленные изменения, и нестабильной, если этот сигнал изменяется внезапно.
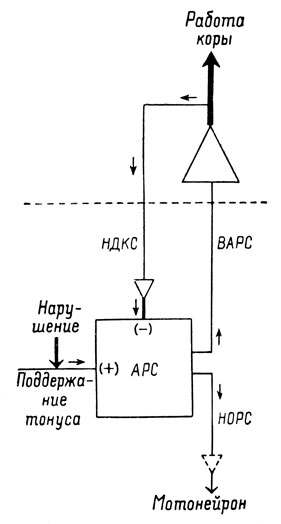
Рис. 7. Схема ретикуло-кортико-ретикулярной цепи. Граница между ретикулярной формацией и корой обозначена пунктирной линией. Размеры изображений соответствуют степени усиления сообщений на уровне различных структур, а толщина линий - их относительной интенсивности в различных частях цепи. АРС - активирующая ретикулярная система; ВАРС - восходящая активирующая ретикулярная система; НДКС - нисходящая дезактивирующая корковая система; НОРС - нисуходящая облегчающая ретикулярная система. (По Южлену А. и Бонвалле М.)
В тех случаях, когда внезапное изменение относится не к увеличению интенсивности стимуляции, а к ее прекращению (условие, часто достаточное, чтобы вызвать пробуждение), мы можем предположить, что действуют аналогичные физиологические механизмы. В самом деле, при прекращении действия какого-либо стимула относительно новую интенсивность по контрасту (к открытию механизмов которого начинает приближаться нейрофизиология) внезапно приобретают другие элементы стиму-ляционного комплекса (см. ниже).
В. Значение
Аффективный или мотивационный "заряд" стимулов имеет, разумеется, первостепенное значение для пробуждения, и неслучайно в этой связи всегда приводятся избитые примеры (мать, пробуждаемая вздохом ребенка и не пробуждаемая шумом, и т. д.). Однако эта проблема значения пробуждающих стимулов ставит психофизиолога перед реальным парадоксом. В самом деле, каким образом нервная система, падение активности которой и приводит к потере критической реактивности, специфичной для сна (Пьерон, 1913), способна различать стимул, до того, как он вызовет ее пробуждение? Эта трудная проблема различительной способности во время сна может дать повод для многих гипотез. Но все эти гипотезы должны исходить из того, что, хотя во время сна активность нервной системы значительно изменяется, она не прекращается (существование сновидений является старым аргументом в пользу этого) и остается чувствительной к изменениям внешнего мира. Поскольку потеря сознания характерна для сна, считается, что высшие энцефалические структуры "отключены" от внешнего мира. В этом случае следует предположить, что сохраняется нечто вроде "бодрствования", обеспечиваемого нижележащими центрами, и допустить существование различительной сенсорной способности на уровне этих центров.
Поскольку интеграция внутренних стимулов, связанных с регуляцией основных функций, осуществляется на подкорковых уровнях, нетрудно понять устойчивость их влияний и их действие на системы пробуждения.
Внешние стимулы, напротив, получают почти все свое значение из приобретенного опыта: это значит, что проблема различения пробуждающих стимулов сводится к проблеме сохранения условных рефлексов во время сна. Несмотря на многочисленные экспериментальные попытки и рекламу некоторых магнитофонных фирм, основанную на вере в "гипнопедическое" обучение, в свое время предложенное Хаксли в книге "Лучший из миров", возможность выработки условных рефлексов во время сна еще не доказана, исключая очень неглубокие стадии сна (см. выводы Эммонса и Саймона, 1955; Саймон, 1964) или случаи, сомнительные в отношении непрерывности состояния сна. Так Бен и Барат (1965) недавно установили, что звук, ассоциированный с электрическим ударом во время глубокого сна у человека, становится способным условнорефлекторно вызывать электрокортикальный ответ (К-комплекс) и что тот же самый звук, исключая звуки близкой частоты, вызывал при возвращении к состоянию бодрствования реакцию блокады α-ритма. Однако возможно, что наличие К-комплекса свидетельствует об очень коротком пробуждении; в таком случае связь выработалась вне настоящего сна. Именно эту гипотезу предложили Изкьердо и сотр. (1965); в очень остроумном эксперименте они вырабатывали условный рефлекс у кошки, используя реакцию пробуждения ЭЭГ в качестве инструментального ответа, позволяющего избежать удара электрического тока во время сна. Эти авторы считают, что подобная связь может образоваться только в периоды пробуждения, вызванные предъявлением подкрепляющего агента.
Зато мы имеем многочисленные экспериментальные доказательства сохранения во время сна условных рефлексов, приобретенных в состоянии бодрствования (В неопубликованном эксперименте, который мы провели в 1953 г. вместе с Ф. Брессоном, нам не удалось обнаружить с помощью метода повторного заучивания какого-либо доказательства возможности обучения во сне. Зато мы вынуждены были вскоре прервать работу, так как испытуемые после нескольких ночей эксперимента систематически просыпались при предъявлении материала для запоминания, хотя последний был перемешан с нейтральными стимулами такой же интенсивности. Этот материал приобрел, следовательно, в ходе последовательных пробуждений особое значение, которое сохранялось и во время сна). Это сохранение проявляется в том, что стимул, получивший условное значение, во сне остается способным вызывать:
а) либо активацию (переход к стадии менее глубокого сна или к полному пробуждению), нечто вроде остаточного компонента первоначальной реакции. Так, Раулэнд (1957) получил у кошки избирательное пробуждение на звук, ранее связанный с электрическим ударом. Освальд, Тэйлор и Трейсмен (1960) показали, что у человека пробуждение легче вызвать, называя собственные имена (и особенно имя испытуемого), на которые по инструкции, данной накануне, следовало отвечать движением руки;
б) либо первоначальный условный ответ без признаков пробуждения. Так, Гранде и Хаммашу (1961) удалось получить у людей инструментальные реакции различения (нажать на ключ, чтобы избежать электрического удара) даже при очень глубоких электроэнцефалографических стадиях сна. Но такие наблюдения редки.
Как бы то ни было, все эти наблюдения имеют два важных следствия: одно - в плане изучения феноменов восприятия, а другое - в отношении изучения условных рефлексов.
Действительно, следует допустить, что во сне сохраняется какая-то различительная перцептивная способность.
С другой стороны, существует возможность активации условных следов. В настоящее время трудно, учитывая уровень наших знаний в отношении сна, связать эти факты с деятельностью той или иной нервной структуры. Однако можно высказать две гипотезы: либо условный след устанавливается на уровне коры, как это всегда утверждали И. П. Павлов и его школа, и в таком случае следует предположить, что спящая кора сохраняет различительную способность и остается способной активировать ретикулярные структуры, которые вызывают ее собственное пробуждение. Либо следует предположить, что местом условных следов являются и подкорковые структуры, что означает также, что различение может осуществляться и на этом уровне. Экспериментальные данные весьма фрагментарны, но они, по-видимому, говорят в пользу последней гипотезы.
Если допустить, что "привыкание", то есть угасание ранее существовавшего ответа, составляет самую примитивную форму научения, то классические эксперименты Шарплеса и Джаспера (1956) действительно показывают, что во время сна возможна дифференциация стимулов, даже после обширного разрушения коры. Эти авторы, получив угасание реакции пробуждения у кошек на звуки определенной частоты, повторяемые одинаково, без подкрепления, показали, что всякое предъявление звука другой частоты немедленно вызывает пробуждение. Удаление значительных участков коры, в том числе полное удаление слуховых зон, не мешало дифференциации звуков "привычных" и впервые предъявляемых.
Однако после подобных разрушений коры оказывается невозможной дифференциация тональных мелодий.
Если вспомнить работы Даймонда и Неффа (1957), показавших, что у бодрствующего животного простое тональное различение сохраняется после разрушения слуховых зон, тогда как для различения мелодий необходима сохранность коры, то из предыдущих экспериментов можно сделать вывод, что простые различительные процессы, несомненно, осуществляются одними и теми же структурами и во сне и при бодрствовании.
Хотя психофизиологические данные не позволяют сделать окончательных выводов относительно роли подкорковых структур в образовании условных рефлексов, по-видимому, установлено, что условные рефлексы могут сохраняться и после полного разрушения коры. Так, Загер (см. Фессар, 1958) обнаружил сохранение "вегетативно-аффективного" компонента (двигательное возбуждение, расширение зрачка и т. д.) защитного условного рефлекса у собаки, предварительно выработанного на определенный стимул. Таким образом, при отсутствии коры эти животные еще способны были воспринимать значение стимула. Однако мы снова должны подчеркнуть, что эта различительная способность, по-видимому, тесно связана со стимулами, имеющими большой мотивационнын "заряд", и реакциями типа эмоционального пробуждения: действительно, большинство опытов относится к сигналам, связанным с электрическим ударом или с потребностью в пище (у животных). Это значит, что во время сна сохраняется аффективное значение стимулов, связанных с основными потребностями. И это снова говорит о близости механизмов пробуждения к более общим процессам активации.
Следует, наконец, отметить, что, по данным Буэндиа и сотр. (1959), степень генерализации стимулов, вызывающих пробуждение, свидетельствует о том, что во сне способность различения слабее, чем в состоянии бодрствования (В нашей лаборатории проводятся в настоящее время эксперименты, ставящие своей целью уточнить этот момент и проследить сохранение значения условных стимулов в зависимости от уровней глубины сна). Имеет ли эта подкорковая различительная способность ту же природу, что и в некоторых простых опытах с подпороговыми сигналами? В самом деле, известно, что вегетативные реакции (например, электрокожная реакция) у человека могут быть вызваны подпороговыми стимулами, предварительно условнорефлекторно связанными на надпороговом уровне (Лазарус и Мак-Клири, 1951).
В настоящий момент мы не располагаем никакими данными, чтобы ответить на этот вопрос, однако следует отметить, что и в этом типе экспериментов действенными остаются стимулы с сильным аффективным "зарядом", даже когда они предъявляются в условиях, не допускающих их вербального узнавания.
В заключение скажем, что способность различения стимулов с тревожным значением сохраняется и при отсутствии их осознания, будь то в период, когда организм находится в состоянии сна или когда стимулы являются подпороговыми. Возможно, что такое различение, менее тонкое, чем в состоянии бодрствования, совершается на уровне ретикулярных структур, но в конечном счете именно ретикулярная активация, вызываемая этими стимулами, влечет за собой соматическое, вегетативное и кортикальное пробуждение.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://psychologylib.ru/ 'Библиотека по психологии'